Phytochrom
Phytochrome sind eine Klasse von weitverbreiteten Photorezeptor-Proteinen, die in Pflanzen, Algen, Bakterien, Cyanobakterien und Pilzen vorkommen. Sie messen das Verhältnis von hellrotem zu dunkelrotem Licht und steuern ein breites Spektrum von Antworten auf Lichtreize, wie etwa die Ergrünung von Pflanzenteilen, die Schattenflucht oder die Samenkeimung bei Pflanzen. Neben den Cryptochromen und den Phototropinen sind sie die wichtigste Klasse an Photorezeptoren.
Es wurde in den 1950er-Jahren durch ein Team um Harry Borthwick entdeckt.
Bedeutung
In höheren Pflanzen steuern Phytochrome eine Vielzahl von Prozessen, unter anderem der Keimung, Photomorphogenese der Keimlinge, Blütenbildung, Photoperiodismus, und Vermeidung von Grünschatten, wie er unter Blattdecken anzutreffen ist. Bei niederen Pflanzen wie bspw. Moosen sind Phytochrome für den Phototropismus und den Polarotropismus verantwortlich.
In Cyanobakterien sind Phytochrome für die chromatische Adaptation des Photosyntheseapparates verantwortlich: die Zusammensetzung der akzessorischen Photosynthesepigmente wird an die spektralen Eigenschaften des Lichts angepasst. Auch in Bakterien, die anoxygene Photosynthese betreiben, wird so die Zusammensetzung des Photosyntheseapparates bestimmt.
In einigen Pilzen entscheidet das Phytochrom-Signal über den Eintritt in die vegetative oder generative Entwicklungsphase.
Reaktion auf Licht: Photokonversion
Phytochrome können in zwei verschiedenen Konformationen vorliegen: die Pr-Form (r für rot, englisch red) besitzt ein Absorptionsmaximum bei 660 nm, die Pfr-Form (fr für dunkelrot, englisch far red) bei 730 nm.[1] Diese spektralen Eigenschaften werden sowohl durch den Chromophor, als auch durch das umgebende Protein bestimmt.
Schon früh wurde erkannt, dass sich die Absorptionseigenschaften des Moleküls und auch die pflanzlichen Antworten durch Bestrahlung mit bestimmtem Licht verändern lassen: im Dunkeln gewachsene Gewebe enthalten ausschließlich die physiologisch inaktive Pr-Form des Phytochroms, das nach Bestrahlung mit hellrotem Licht in die aktive Pfr-Form übergeht. Die Pflanze zeigt daraufhin typische Reaktionen wie zum Beispiel Photomorphogenese. Durch Bestrahlung mit dunkelrotem Licht kann dieser Effekt rückgängig gemacht werden – Pfr geht wieder in die inaktive Pr-Form über. Pfr kehrt durch seine thermodynamische Instabilität lichtunabhängig wieder in die Pr-Form zurück. Dieser Prozess wird Dunkelreversion oder Dunkelkonversion genannt.
Die biologischen Antworten werden so durch das Verhältnis Pr/Pfr und das Ausmaß der Dunkelreversion bestimmt.
Dieser Prozess wird durch eine Konformationsänderung des Chromophors bei Bestrahlung mit entsprechendem Licht veranlasst, die eine Strukturänderung des gesamten Proteins nach sich zieht. Dabei wird die Kinase-Aktivität des Proteins verändert. In Pflanzen werden die weit voneinander entfernten Domänen P3/GAF und PAS-A und PAS-B exponiert. Sie tragen Signale für den Transport in den Kern und möglicherweise Interaktionsflächen für Partnerproteine.
Proteinaufbau
Phytochrome sind in weiten Teilen stark konserviert, allen ist eine N-terminale Sensorregion und eine C-terminale Regulator- bzw. Dimerisierungsregion gemein.
Die Sensorregion ist aus den drei Domänen P2/PAS, P3/GAF und P4/PHY aufgebaut, wobei das Chromophor bei Bakterien an der P2/PAS und bei Pflanzen an der P3/GAF-Domäne mit dem A-Ring kovalent über eine Thioesterbindung an ein Cystein des Proteins gebunden wird und in eine tiefe Tasche eingesenkt ist. Bei Pflanzen gibt es des Weiteren eine zusätzliche N-terminale P1-Domäne, die in den verschiedenen Arten unterschiedliche Funktionen hat. P2/PAS und P3/GAF sind essentiell für die Lichtwahrnehmung und die Signaltransduktion besitzen Bilin-Lyase-Aktivität, die für den Einbau des Chromophors erforderlich ist. P4/PHY ist für die spektralen Eigenschaften, die Kinase-Aktivität und die Feinabstimmung der Aktivität des Phytochroms verantwortlich und veranlasst in Pflanzen den Transport in den Kern.
Die Regulatorregion besitzt in der ursprünglichen Form eine Domäne mit Histidinkinase-Aktivität, mit deren Hilfe Proteine phosphoryliert werden können. In Pflanzen ist diese Funktion verloren gegangen, bei ihnen wird die Kinase-Aktivität durch eine Serin-Threonin-Kinase bewerkstelligt, die jedoch im N-Terminus des Proteins gefunden wurde. Pflanzen besitzen zusätzliche PAS-A und PAS-B-Domänen, die für die Dimerisierung und den Transport in den Kern verantwortlich sind.
In Eukaryoten treten Phytochrome immer als Dimere auf, in Prokaryoten nur als Monomer.
Struktur des Chromophors bei Pflanzen
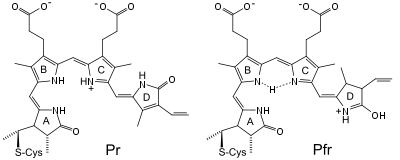
Je nach Organismengruppe sind verschiedene Chromophore anzutreffen, die vom Häm abgeleitet sind. Bakterien und Pilze verwenden Biliverdin IX α, höheren Pflanzen Phytochromobilin, Cyanobakterien und Algen Phycocyanobilin.
Das Chromophor ist ein lineares Tetrapyrrol, das an den Ringen A,B und C durch seine Umgebung fixiert ist. Infolge der Photokonversion kann der D-Ring nach Aufnahme eines hellroten Lichtquants rotieren. Das Molekül geht so in eine cis-trans-Isomer Form an der Doppelbindung zwischen C15 und C16 über. Die Pr-Form des Moleküls ist C15-Z,anti-konfiguriert, die Pfr-Form besitzt C15-E,anti-Konfiguration. Die Konformationsänderung des Chromophors zieht eine Konformationsänderung des gesamten Proteins nach sich, das dadurch von Pr- in die Pfr-Form übergeht. Dieser Vorgang wird durch Bestrahlung mit dunkelrotem Licht umgekehrt.
Genfamilie der pflanzlichen Phytochrome
In Arabidopsis thaliana wurden bislang fünf Gene für Phytochrome gefunden, drei in Reis, vier in Kiefer und drei in Ginkgo biloba. Phylogenetische Analysen haben ergeben, dass die Phytochrome sich vor der Divergenz der Samenpflanzen in die zwei Hauptgruppen Phy A und Phy B aufgespalten haben, in die sich alle Phytochrome einordnen lassen. Bei Ceratodon purpureus, einem Moos, sind bisher 4 Phytochrome bekannt, eines davon weist eine für Phytochrome bisher einmalige Gensequenz auf, jedoch konnte bisher nicht geklärt werden, ob es sich bei dieser Version um ein funktionelle Protein oder um ein Pseudogen handelt.
Phy A kommt fast ausschließlich in großen Mengen in etiolierten Pflanzen vor, da seine Pfr-Form instabil ist und seine eigene Gentranskription hemmt. Phy B kommt in unter normalen Lichtbedingungen gezogenen Pflanzen vor, sowie in kleinen Mengen in etiolierten Pflanzen. Die Pfr-Form von Phy B ist im Gegensatz zu Phy A stabil und wird konstitutiv exprimiert.
Viele der Phytochrome haben ähnliche Eigenschaften und teilen sich bestimmte Aufgaben, außerdem haben viele von ihnen spezialisierte Rollen in ihrer biologischen Funktion.
Signaltransduktion
Die intrazelluläre Signaltransduktion ist weitgehend unbekannt. Es ist aber anzunehmen, dass die Histokinase-Domäne in Prokaryoten eine wichtige Rolle in der Übertragung von Phosphatresten spielt. In Eukaryoten liegt keine Histokinase-Aktivität mehr vor, jedoch ist eine Serin-Threonin-Kinase-Aktivität vorhanden.
In Eukaryoten ist eine Translokation des Proteins in den Kern nachgewiesen, wo es Wechselwirkungen mit Transkriptionsfaktoren eingehen und dort direkt die Genexpression beeinflussen kann. Es werden aber auch cytosolische Antworten generiert.
Literatur
- Peter Schopfer, Axel Brennicke: Pflanzenphysiologie. 6. Aufl. Elsevier Spektrum Akademischer Verlag, München 2006, ISBN 978-3-8274-1561-5.
- Nathan C. Rockwell u. a.: Phytochrome Structure and Signaling Mechanisms. In: Annual Review of Plant Biology, Jg. 57 (2006), S. 837–58, ISSN 1543-5008.
- Kurt Schaffner: Zur Photophysik und Photochemie von Phytochrom, einem photomorphogenen Regler in grünen Pflanzen, in: Rheinisch-Westfälische Akademie der Wissenschaften (Natur-,Ingenieur- und Wirtschaftswissenschaften, Vorträge), vol 362, Opladen : Westdeutscher Verlag 1988, ISBN 978-3-531-08362-9.
Einzelnachweise
- Phytochrome. In: Spektrum der Wissenschaft, Spektrum Akademischer Verlag, Heidelberg, 2001.