Mauergecko
Der Mauergecko (Tarentola mauritanica) ist ein häufiger und im Mittelmeerraum großflächig verbreiteter, nachtaktiver Gecko. Er gehört zur Gattung Tarentola, innerhalb der Familie der Blattfingergeckos (Phyllodactylidae).
| Mauergecko | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Mauergecko (Tarentola mauritanica) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Tarentola mauritanica | ||||||||||||
| (Linnaeus, 1758) |

Merkmale
Der Mauergecko zählt zu den größten Geckoarten in Europa. Er erreicht eine Körperlänge von bis zu 16 cm, wobei die Kopf-Rumpf-Länge zwischen 74 und 84 mm liegt.[1][2][3] Männchen werden größer als die Weibchen und besitzen deutlich breitere Köpfe. Die Grundfarbe der Mauergeckos besteht gewöhnlich aus grauen und bräunlichen Tönen. Meistens finden sich weitere hellere Flecken, sowie dunkle Querstreifen (vier bis fünf zwischen Nacken und Sakralregion.[2]) auf dem Körper, die jedoch mit dem Alter verschwinden [1] Der Körper ist typischerweise abgeflacht, die Schnauze breit und spitz zulaufend. Femoral- oder Preanalporen (Hautdrüsen) sind bei keinem der beiden Geschlechter zu finden[1] Jedoch wurde auch schon beschrieben, dass sich bei Männchen im Alter von 2 bis 3 Jahren unterhalb der Kloake zwei markante Hautöffnungen (vermutlich Drüsenöffnungen) befinden, die bei den Weibchen nicht vorhanden sind und zur Geschlechtsbestimmung hilfreich seien können [4]
Die Augen haben eine senkrecht geschlitzte Pupille, die Iris ist grau gefärbt. Am Kopf, Körper, Schwanz und Beinen befinden sich ausgeprägte Tuberkelschuppen, die in parallelen Reihen angeordnet sind. Am Körper befinden sich quer 12 bis 16 Reihen.[2] An der Grenze zwischen den Bauch- und Rückschuppen sind allerdings keine vergrößerten Tuberkel zu finden. Die Rückentuberkel sind mehrfach gekielt und mit Sensillen besetzt.[2] Dabei kann der Kiel der Tuberkelschuppen innerartlich unterschiedlich spitz ausgeprägt sein, die Tuberkel sind jedoch nie nach caudal gerichtet.[2] Um die Tuberkel befinden sich hufeisenförmige Rosetten aus Schuppen intermediärer Größe (Körnerschuppen), die restlichen Rückenschuppen sind hingegen deutlich größer [2] Die Schuppen von regenerierten Schwänzen sind deutlich kleiner, als die des restlichen Körpers [5], wodurch ein Schwanzregenerat vom Körper abgesetzt wirkt und dadurch schnell erkennbar ist. Die Zehen des Mauergeckos sind abgeflacht und besitzen am dritten und vierten Finger deutlich sichtbare Krallen. Weibchen besitzen auch an den restlichen Zehen Krallen, die jedoch kaum sichtbar und häufig verdeckt sind[4] An der Unterseite befinden sich bei allen Zehen ungeteilte, breite Haftlamellen.
Haftstrukturen
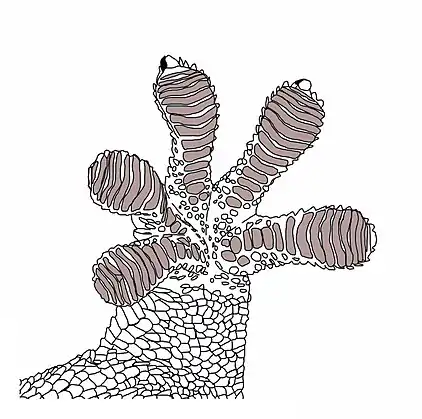
Der Mauergecko besitzt an der Sohle seiner Zehen spezielle Haftstrukturen, die es ihm ermöglichen, selbst an sehr glatten Oberflächen entlang zu klettern. Diese Strukturen sind lamellenartig angeordnet. Unter dem ersten Zeh befinden sich rund 11, unter dem vierten rund 16 und unter dem fünften rund 20 Lamellen.[3] Jede Lamelle besteht aus Tausenden von Setae (dünne, haarartige Strukturen), die sich wiederum in eine große Anzahl von tellerförmig verbreiterten Spatulae (Hafthärchen) aufgliedern.[6] Beim Mauergecko ist eine Seta rund 150 µm lang [7] Die Spatulae, als letzte Struktur des hierarchisch gegliederten Haftsystems, bilden den Kontaktpunkt zur Oberfläche. Zwischen diesen beiden Punkten wirken „van der Waals-Kräfte“ (molekulare Adhäsion), die verantwortlich für die Haftfähigkeit der Geckos sind [8]
Die Entwicklung der Haftstrukturen wurde beim Mauergecko durch Hiller (1972) näher untersucht. Bereits nach einer Häutung wird die übernächste Epidermisgeneration und damit auch die Haftstrukturen angelegt. Dies erfolgt vor allem in der Hellen Schicht und der Oberhäutchenzellen-Schicht. Als erster Schritt erfolgt die Aufspaltung der Setae-Enden. Dies geschieht durch das Einwachsen von Keratinfilamentbündeln von der Oberhäutchenzellen-Schicht in die Helle Schicht. Dadurch besitzt die Helle Schicht eine Matrixfunktion und ist bei der präzisen Ausbildung dieser Strukturen maßgeblich beteiligt. Nach diesem Prozess werden durch gleichzeitiges Auseinanderrücken der Hellen Schicht und der Oberhäutchenzellen-Schicht (Abstand zwischen den Zellschichten vergrößert sich langsam) sowie weiterer Keratinisierung die eigentlichen Haftstrukturen gebildet. Die Helle Schicht behält bis zur nächsten Häutung ihre Matrixfunktion bei.[7]
Verbreitung
Der Mauergecko kommt großflächig in den Küstenregionen des mediterranen Raums vor. Neben Portugal,[9][10] Spanien,[11] Gibraltar,[12] Frankreich (inklusive Korsika),[13] Italien,[14][15] (inklusive Sardinien[16][17] und Lampedusa), den Balearischen Inseln (eingeschleppt in Mallorca, Menorca, Ibiza, Formentera und Cabrera [18]), Malta, den Azoren [19] den Küstenregionen Kroatiens (außer Istrien) und den adriatischen Inseln, werden auch Teile Nordafrikas besiedelt. Dazu zählen Marokko,[20] Algerien,[21] Tunesien,[22] Libyen, Ägypten, Westsahara,[23] Israel und der Sinai.[24]
Es ist davon auszugehen, dass die Vorkommen im Osten des mediterranen Raums auf Verschleppung durch den Menschen zurückzuführen sind.[1] So auch in Griechenland, wo die Tiere auf dem Festland nur auf der Peloponnes vorkommen (rund um die Hafenstädte Patras[5] und Pylos.[25] Darüber hinaus auf den vorgelagerten Inseln Kephallonia, Ithaka, Zakynthos, Kreta, Strofades, Dia,[1] Korfu[26] und Lesvos [27] Auf den restlichen Inseln der Ägäis ist der Mauergecko nicht verbreitet[5] Auch das Vorkommen der Inselgruppe Madeira geht auf anthropogene Einführung zurück [28][29]
Außerhalb Europas wurde der Mauergecko auch nach Uruguay,[30][31] in die USA (Kalifornien und San Diego)[27], Argentinien[31] sowie Chile (Santiago de Chile)[32] verschleppt, wo er sich ebenfalls erfolgreich ansiedelte.
Lebensraum
Der Mauergecko bevorzugt trockene und steinige Habitate, wie Klippen, Steinmauern und Ruinen. Selbst in der Nähe menschlicher Siedlungen ist er häufig zu finden. Dort hat er sich als äußerst erfolgreicher Kulturfolger etabliert, der gerne an Häuserfassaden selbst inmitten einer Stadt entlang klettert. Bevorzugt werden dort vor allem alte Gemäuer mit vielen Rissen und Spalten, die den Tieren als Unterschlupf und Versteck dienen. Von einem sehr ungewöhnlichen Tagesunterschlupf berichtet Barts (2013), der einen adulten Mauergecko in einem Wespennest fand.[33] Tlili et al. (2012) konnten Mauergeckos an Elektrizitätsmasten, Ruinen, kleinen Autobrücken, Landmarken und selbst in der unmittelbaren Nähe von Müll finden.[34] Besonders zahlreich kann der Mauergecko in leerstehenden Häusern (Innen- und Außenwänden) und an alten Brunnen beobachtet werden.[5] In der Nacht findet man ihn darüber hinaus regelmäßig in der Nähe von künstlichen Lichtquellen, da er dort nach angelockten Insekten jagt. Im Allgemeinen gilt der Mauergecko innerhalb der Gattung Tarentola als am stärksten anthropogen-assoziierte Spezies [34]
Während der Mauergecko an einigen Punkten des Verbreitungsgebietes ausschließlich auf die Küstenregion beschränkt ist (bspw. Tunesien.[34], Griechenland.[5]), kann er auf der Iberischen Halbinsel auch im Inland angetroffen werden [2] Dort bewohnt er Biotope von Meereshöhe bis etwa 2.300 m (in Spanien).[5]

Unterarten
Der Mauergecko wurde 1758 von Carl von Linné als Lacerta mauritanica erstbeschrieben. Die Terra typica umfasst „Mauritantia“, also das nördliche Afrika. Linné erhielt das Exemplar vom schwedischen Konsul in Algier.,[35] die Typuslokalität beschränkt sich damit auf Algier.[2]
Es werden derzeit 3 Unterarten unterschieden:
Tarentola mauritanica mauritanica (Linnaeus 1758)
Tarentola mauritanica juliae (Joger 1984)[2]
Tarentola mauritanica pallida (Geniez et al. 1999)
Die Nominatform Tarentola mauritanica mauritanica ist in den europäischen Küstenregionen des Mittelmeers (auf der Iberischen Halbinsel auch im Landesinneren) einschließlich zahlreicher Mittelmeerinseln (u. a. eines Teils der Ionischen Inseln und aller größeren Inseln des westlichen Mittelmeeres) verbreitet. Darüber hinaus kommt sie auch in Marokko nördlich des Hohen Atlas, in Algerien nördlich des Sahara-Atlas und in Tunesien nördlich der Linie Gafsa–Sfax vor.[2]
Die Terra typica der Unterart Tarentola mauritanica juliae umfasst den östlichen Antialtias in Marokko. Tarentola m. juliae ist in Südmarokko verbreitet. Dort kann die Unterart des Mauergeckos im Bereich des westlichen Hohen Atlas und seiner Ausläufer, sowie südlich bis zum Hauptkamm des Antiatlas und seiner Ausläufer angetroffen werden.[2] Der Holotypus ist mit einer gedrungenen Gestalt mit kurzem Kopf und dünnem Schwanz beschrieben worden. Die Rückentuberkel sind klein und spitz, aber nur wenig hervorgehend. Rosetten um die Tuberkel sind vorhanden.[2] Im Norden des Verbreitungsgebietes gibt es Intergradationszonen mit Übergangsformen zu Tarentola m. mauritanica [2]
Das Verbreitungsgebiet der Unterart Tarentola mauritanica pallida ist auf den atlantischen Randbereich von Südmarokko und Westsahara von der Mündung des Oued Drâa bis Dchira beschränkt.[36] Tarentola m. juliae und Tarentola m. mauritanica kommen ebenfalls in diesem Gebiet vor. Tarentola m. pallida unterscheidet sich jedoch von diesen durch eine rosafarbene bis gelbliche Grundfärbung und ein durch glatteres, weniger warziges Erscheinungsbild (flachere Tuberkelschuppen auf der Rückenmitte) [36]
Die im östlichen Nordafrika verbreitete Art Tarentola fascicularis (Daudin 1802) wurde früher ebenfalls als Unterart des Mauergeckos betrachtet, aber 2010 als eigene Art anerkannt.[3]
Taxonomie
Durch Untersuchungen der mitochondrialen DNA konnten sechs genetisch unterschiedliche Linien entlang Nordafrikas nachgewiesen werden. Diese Phylogruppen erstrecken sich offenbar in Algerien, Tunesien, Libyen, in dem Süden Marokkos, in dem Norden Marokkos und in Zentralmarokko. Im Gegensatz dazu werden die Populationen entlang Spaniens, Portugals, Italiens, Kretas, Menorcas und Tunesiens durch bloß einen einzigen Haplotyp charakterisiert. Es ist daher davon auszugehen, dass diese Populationen auf eine anthropogene Einführung mit anschließend rascher Ausbreitung entlang des europäischen Mittelmeerraumes zurückzuführen sind.[37] Ihre Ursprünge finden sich möglicherweise in Marokko.[38]
Die hohe genetische Diversität des Mauergeckos insbesondere in Nordafrika könnte dafür sprechen, dass Tarentola mauritanica einen ganzen Arten-Komplex repräsentiert.[37]
Ernährung
Der Mauergecko ist ein opportunistischer Räuber, der ein breites Nahrungsspektrum besitzt. Das Nahrungsspektrum besteht vor allem aus Gliederfüßer, wie Spinnen, Käfer, Fliegen, Ameisen und Zikaden.[39] Darüber hinaus konnten in Mägen Pflanzensamen, Schalenreste von Geckoeiern und Häutungsreste gefunden werden.[39] Die Nahrungszusammensetzung, als auch das Jagdverhalten kann sich jedoch mit dem jeweilig besiedelten Habitat unterscheiden. Mauergeckos, die natürliche, zumeist aride und felsige Lebensräume besiedeln, haben sich auf bodenbewohnende Arthropoden spezialisiert [40] Dies deckt sich auch mit dem Verhalten anderer Geckoarten aus ariden Gebieten.[41] Aufgrund des geringen Nahrungsangebots solcher Lebensräume, müssen die Mauergeckos aktiv nach Nahrung suchen („foraging“). Diese Verhaltensweise kann jedoch eingeschränkt sein, da die Umgebungstemperatur in der Nacht zum Teil stark abkühlt.[40] Ein anderes Verhalten zeigen Mauergeckos, die in urbanen Gebieten vorkommen. Gerne werden von diesen in der Nacht künstliche Lichtquellen aufgesucht, da sich dort angelockte Fluginsekten befinden. Die Nahrung kann bis zu 24 % aus Insekten der Ordnung Diptera und adulten Lepidoptera bestehen [40] Mauergeckos verharren häufig an den Lichtquellen und warten auf die Beute, ohne aktiv danach zu suchen („sit and wait“). Weiterhin herrscht in Städten eine niedrigere, nächtliche Abkühlung, was der Aktivität der Mauergeckos förderlich zu seien scheint [40]
Gil et al. (1993) konnten auch Unterschiede im Nahrungsspektrum zwischen Festland- und Inselbewohnern feststellen. Da auf Inseln häufig eine niedrigere Beutediversität besteht, verlagert sich die Ernährung auf wenige, häufig auftretende Beutetypen (in diesem Fall vor allem Käfer). Auf dem Festland ist hingegen die Diversität groß, sodass auch die Nahrung vielseitig zusammengesetzt seien kann.[39]
Neben Arthropoden werden vom Mauergecko auch andere Echsen und sogar Artgenossen gejagt und gefressen.[5][40] Kleinere Geckoarten und Artgenossen werden dadurch schnell aus dem Revier der Mauergeckos verdrängt.[42] Es ist daher anzunehmen, dass der Mauergecko als invasive Art fremde Fauna stark negativ beeinflussen könnte (Artenverdrängung) [27]
Verhalten
Die Hauptaktivität des Mauergeckos liegt in den Dämmerungs- und Nachtstunden. Dann jagen sie entweder aktiv oder lauernd nach Nahrung. Begrenzt wird nachts die Aktivitäts- und damit auch Jagddauer von der sinkenden Körpertemperatur. Gil et al. (1993) gehen davon aus, dass sich damit auch das Jagdverhalten saisonbedingt ändert.[39] So zeigen die Mauergeckos während der kühlen Perioden ein verstärktes „Sit-and-wait“-Verhalten, mit welchem sie unselektiv Beute machen. In den Sommermonaten hingegen ist die Aktivität der Tiere deutlich erhöht, sodass nun aktiv und selektiv nach Beutetieren gejagt werden kann.
Auch am Tage kann der Mauergecko zuweilen beim Sonnenbad beobachtet werden. Dann befindet er sich meist nie mehr als 2 bis 4 cm von seinem Unterschlupf (meist Felsspalte) entfernt und flüchtet unverzüglich in diesen, sobald sich eine Bedrohung nähert.[43] Die meiste Zeit der Tagesaktivität wird mit der Thermoregulation verbracht, kaum mit territorialen Auseinandersetzungen. Dennoch herrscht stets ein großer Individualabstand zwischen gleich großen Adulti (häufig 2 bis 3 m).[43] Während des Sonnenbades verändern die Mauergeckos ihre Hautfarbe zu dunklen Braun- oder Schwarztönen, um so viel wie möglich des Sonnenlichtes zu absorbieren. Im Terrarium liegt die bevorzugte Körpertemperatur des Mauergeckos bei 31,6 °C.,[44] in situ wurden Körpertemperaturen von rund 25 °C gemessen[45] An bewölkten oder regnerischen Tagen ist der Mauergecko tagsüber nicht aktiv und versteckt sich in seinem Unterschlupf [43]
Werden Mauergeckos ergriffen, zum Beispiel durch einen Menschen, so zeigen sie verschiedene Abwehrverhaltensweisen. Neben Körperwinden, Drohen mit offenem Maul, Koten und quiekenden Lauten, versuchen sie sich durch kräftige Bisse zu verteidigen.[43]

Schwanzautotomie
Der Mauergecko ist in der Lage seinen Schwanz abzuwerfen (Autotomie). Der Abwurf des Schwanzes erfolgt bei Bedrohung durch einen Räuber oder bei intraspezifischen Interaktionen. Der Beutegreifer soll durch den Schwanz irritiert werden, sodass der Mauergecko fliehen kann. Der Verlust des Schwanzes kann sich jedoch auf die Lokomotorik des Geckos, die Energiereserven und auf die Fähigkeit sich erfolgreich zu paaren negativ auswirken.[46][47] Regenerierte Schwänze wachsen kegelförmig nach und sind von kleinen Schuppen besetzt.[5] Des Weiteren sind sie kürzer und massiger als das Original [46] Bei den Unterarten Tarentola m. mauritanica und Tarentola m. juliae besitzen die Regenerate eine graue Färbung und eine schwärzliche, längsgerichtete Marmorierung. Die Regenerate der Unterart Tarentola m. pallida weisen hingegen eine gelbe Färbung auf, sowie eine rankenförmige, purpurne Marmorierung.[48] Die prozentuale Zusammensetzung an Lipiden, Proteinen und Asche unterscheidet sich zwischen dem Originalschwanz und dem Regenerat[46] So ist im Regenerat des Mauergeckos der prozentuale Anteil von Proteinen und Asche leicht niedriger als im Originalschwanz, der Anteil an Lipiden ist jedoch annähernd doppelt so hoch. Dies deckt sich auch mit den Daten anderer Geckoarten[46] und unterstützt die Hypothese über den adaptiven und funktionalen Wert der Reproduktion des Schwanzregenrates bei Geckos (siehe Perez-Mellado et al. 1997).

Rufparameter
Der Mauergecko besitzt ein außerordentlich komplexes Rufrepertoire. Bei Bedrohung, zum Beispiel durch Ergreifen oder auch durch andere Artgenossen, verwendet er vor allem kurze, knurrende Schreie, die allerdings innerartlich sehr variieren können.[49] Gleichzeitig wird dem Angreifer häufig das geöffnete Maul präsentiert. Neben den Abwehrlauten, spielen Rufe auch bei der innerartlichen Kommunikation eine sehr große Rolle, die das Sozialverhalten der Tiere widerspiegeln. Männliche Tiere erzeugen Rufreihen (Einzelrufe kommen fast nie vor), die aus 3 bis 11 Wiederholungen bestehen können. Der erste Ruf der Reihe ist ca. 80 ms lang und besitzt eine Frequenz von bis zu 5 kHz (Hauptintensität zwischen 0 und 4 kHz). Die darauffolgenden Rufe sind kürzer (60 ms) und erreichen eine Frequenz von 4 kHz (Hauptintensität zwischen 0,8 und 1,5 kHz). Die Lautäußerung kann als ein „Tschok…kock…kock…“ beschrieben werden. Der Ruf ist sehr laut (bis 10 m Entfernung hörbar) und wird vor allem mit der Kommunikation mit einem Weibchen innerhalb des Reviers in Verbindung gebracht.[49] Er besitzt also eine werbende und territoriale Funktion. Die Weibchen antworten hierauf mit einem Einzelruf, der einem „Keck“ oder „Kjäck“ ähnelt. Dieser Ruf kommt nur während der Paarungszeit vor, wird jedoch auch von paarungsunwilligen Weibchen geäußert, sodass er nicht im Allgemeinen Paarungsbereitschaft signalisiert. Der Ruf ist deutlich höherfrequenter, als der der Männchen und erreicht bis zu 10 kHz (Hauptintensität zwischen 1,2 und 5 kHz). Die Dauer beträgt rund 70 ms [49] Auch Jungtiere zeigen vereinzelt Vokalisierungen. Zwei Einzelrufe können hierbei unterschieden werden. Zum einen ein „Klik“ – Ruf (20 ms lang, bis 16 kHz, Hauptintensität zwischen 6 und 10 kHz) und zum anderen ein „Krek“ – Laut (50 ms lang, bis 9 kHz, Hauptintensität zwischen 3 und 8 kHz). Diese Rufe treten sowohl in der Gruppe, als auch bei vereinzelten Tieren auf [49]
Fortpflanzung
Mauergeckos sind sehr territorial. Während der Paarungszeit kann der Mauergecko häufig paarweise oder in kleineren Gruppen, bestehend aus einem Männchen und mehreren Weibchen, angetroffen werden.[1] Männliche Rivalen werden jedoch aus dem Revier mit quiekenden Lauten und Bissen vertrieben.[5] Eine wichtige Rollte spielt während der Paarungszeit die innerartliche Kommunikation, die über ein reiches Spektrum verschiedener Vokalisierungen erfolgt. Ist ein Männchen paarungsbereit, patrouilliert es durch sein Revier und beginnt verschiedene Rufreihen.[49] Das Weibchen antwortet darauf mit verschiedenen „Keck“ und „Kjäck“-Rufen. Beim Zusammentreffen berührt das Männchen das Weibchen mit der Zunge. Die Zungenprüfung wird abgewechselt von weiteren Rufen. Anschließend verbeißt sich das Männchen in die Tuberkelschuppen des Weibchens an der Ohrregion und versucht den Schwanz unter dem des Weibchens zu bringen, um damit die Kloaken der beiden Tiere in Kontakt zu bringen. Paarungsbereite Weibchen heben den Schwanz hierfür deutlich an, während paarungsunwillige sich fest an den Bodengrund drücken, um damit eine Paarung zu verhindern [49] Die Kopulationsdauer beträgt zwischen 20 und 60 s. Meistens ändert danach das Männchen die Position, in dem es den Schwanz auf die andere Seite umschlägt, um erneut von dieser Seite zu kopulieren (gleiche Kopulationsdauer). Die Paarung wird durch das Männchen beendet. Häufig kommt es danach zu intensiven Vokalisierungen zwischen den beiden Geschlechtern [49]
Die Weibchen legen abhängig von ihrer Körpergröße mehrmals im Jahr ein bis zwei Eier (zweieiige Gelege werden gewöhnlich ab einer Kopf-Rumpf-Länge von 6 cm gelegt, ungefähres Verhältnis Eineiiges- / Zweieiiges Gelege liegt bei 1 zu 8) zwischen Felsspalten, Holz und Mauerwerk.[49] Auch das Vergraben der Eier im Sand konnte beobachtet werden.[50] Im Terrarium reicht der Eiablagezeitraum von Februar bis August. Es werden innerhalb dieses Zeitraums zwischen 3 und 15 Gelege abgesetzt. Zwischen zwei Eiablagen benötigen die Weibchen lediglich 10 bis 21 Tage. Die Eier haben eine Länge von 1,10 bis 1,30 cm und eine Breite von 0,85 bis 1,10 cm.[49] In freier Natur schlüpfen die Jungtiere, abhängig von der Umgebungstemperatur, nach 40 bis 80 Tagen.[5] Im Terrarium schlüpfen sie bei einer Temperatur zwischen 27 und 30 °C nach 70 bis 75 Tagen. Bei diesen Inkubationsparametern schlüpften bei[49] von 33 Eiern 100 % Weibchen, sodass womöglich von einer temperaturabhängigen Geschlechterbestimmung ausgegangen werden kann. Beim Schlupf sind die Jungtiere zwischen 2,30 und 2,65 cm lang (Kopf-Rumpf-Länge) und besitzen ein Gewicht von 0,26 bis 0,46 g [49] Ab einer Kopf-Rumpf-Länge von 4,5 cm bzw. in einem Alter von 2 bis 3 Jahren gelten die Tiere als paarungsbereit [51][4]

Feinde
Der Mauergecko lebt selbst räuberisch, steht jedoch insbesondere als Jungtier relativ weit unten in der Nahrungskette. Aaskrähen, Kolkraben und Würger bringen regelmäßig Reptilien zu ihren Nestern. Darüber hinaus jagen nachts Katzen, Igel, Steinmarder, Wiesel und Spitzmäuse nach Reptilien.[5] Einige Schlangenarten, wie die Eidechsennatter, Schlanknatter und die nachtaktive Europäische Katzennatter haben sich auf Eidechsen und Geckos spezialisiert und jagen nach diesen zwischen den Mauerspalten.[52] Junge Mauergeckos müssen sich des Weiteren selbst vor ihren eigenen Artgenossen in Acht nehmen.[5] In den ersten Lebenswochen dienen Geckos sogar größeren Insekten und Spinnentieren als Nahrung [5][53]
Gefährdung
Der Mauergecko wird in der IUCN Red List (Version 2019-1) unter „Least Concern“ (LC, niedrigste Bedrohungsstufe) geführt. Lediglich die „ägyptischen Populationen“ seien durch gezieltes Absammeln für den Tierhandel sowie durch Lebensraumverkleinerung bedroht[54] (allerdings handelt es sich dabei wahrscheinlich um die Populationen von Tarentola fascicularis, die von der Redlist anscheinend nach wie vor als Unterart von T. mauritanica ausgehalten wird). Die geringe Bedrohungsstufe ist vor allem mit der großen Verbreitung, der hohen Anpassungsfähigkeit innerhalb der unterschiedlichsten Lebensräume und den starken Populationszahlen begründet. Innerhalb des Verbreitungsgebietes gehört der Mauergecko mit zu den am häufigsten anzutreffenden Reptilien überhaupt.[55][56][15] Die Population wird daher als stabil angesehen. Der Mauergecko ist des Weiteren im Annex III der Berner Konvention, dem Übereinkommen über die Erhaltung der europäischen wildlebenden Pflanzen und Tiere und ihrer natürlichen Lebensräume, gelistet.[1] In Griechenland ist er zudem gesetzlich geschützt (Presidential Decree 67/1981).[1]
Einzelnachweise
- Efstratios D. Valakos, Panayiotis Pafilis, Konstantinos Sotiropoulos, Petros Lymberakis, Panayiota Maragou, Johannes Foufopoulos: The amphibians and reptiles of Greece (= Frankfurter Beiträge zur Naturkunde. Band 23). Edition Chimaira, Frankfurt am Main 2008.
- Ulrich Joger: Taxonomische Revision der Gattung Tarentola (Reptilia: Gekkonidae). In: Bonner Zoologische Beiträge. Band 35, Nr. 1/3, 1984, S. 129–174 (zobodat.at [PDF; 15,8 MB]).
- Ulrich Joger, Ismail Bashaenia: A new Tarentola subspecies (Reptilia: Gekkonidae) endemic to Tunisia. Bonn Zool. Bull. 57 (2). 2010, S. 267–274.
- Arnold, U.: Vereinfachte Geschlechtsbestimmung bei Tarentola delalandii (Duméril & Bibron, 1836) und Tarentola mauritanica (Linnaeus, 1758). Sauria 35 (2). 2013, S. 61–62.
- Trapp, B.: Reptilien und Amphibien des griechischen Festlandes. Natur und Tier Verlag (Münster). 2007 (300 S.).
- Hiller, U.: Morphologische Untersuchungen der Haftborstenbildung und Häutung bei Tarentola mauritanica (Rept.). forma et funetio 2. 1970, S. 169–177.
- Hiller, U.: Licht- und elektronenmikroskopische Untersuchungen zur Haftborstenentwicklung bei Tarentola m. mauritanica L.(Reptilia, Gekkonidae). Zoomorphology, 73(3). 1972, S. 263–278.
- Autumn, K., Liang, Y. A., Hsieh, S. T. & Zesch, W.: Adhesive force of a single gecko foot-hair. In: Nature. Band 405(6787), 2000, S. 681.
- Pfau, J.: Beitrag zur Verbreitung der Herpetofauna in der Niederalgarve (Portugal). Salamandra 24 (4). 1988, S. 258–275.
- Malkmus, R.: Die Amphibien und Reptilien Portugals, Madeiras und der Azoren. Die Neue Brehm-Bücherei, Bd. 621. Westarp Wissenschaften (Magdeburg)/Spektrum Akademischer Verlag (Heidelberg). 1995.
- Busack, S. D., Gabriel, M. B. & Mihalca, A. D.: Tarentola mauritanica (Moorish gecko) eye disease. Herpetological Review 46 (4). 2015, S. 635–636.
- Cortés, J.E.: The herpetofauna of Gibraltar - status, recent history and current research. British Journal of Herpetology 6. 1982, S. 273–275.
- Pottier G., Paumier J.-M., Tessier M., Barascud Y., Talhoët S., Liozon R., D’Andurain P., Vacher J.-P., Barthe L., Heaulmé V., Esslinger M., Arthur C.-P., Calvet A., Maurel C. & Redon H.: Atlas de répartition des reptiles et amphibiens de Midi-Pyrénées. Les atlas naturalistes de Midi-Pyrénées. Nature Midi-Pyrénées, Toulouse. 2008 (126 S.).
- Bruekers, J.: Waarnemingen aan de Ruïnehagedis (Podarcis sicula sicula) en de Muurgeckko (Tarentola mauritanica) in Noord-Italië (Gardameer). Lacerta 64 (3). 2006, S. 101–105.
- Wirth, M.: Toskana. Ein herpetologischer Überblick. Terraria/Elaphe 2013 (3). 2013, S. 41-24.
- Salvi, D., & Bombi, P.: Reptiles of Sardinia: updating the knowledge on their distribution. Acta Herpetologica, 5(2). 2010, S. 161–178.
- Bassu, L., V. Nulchis, M. G. Satta, C. Fresi & Corti C.: Atlas of amphibians and reptiles of Sardinia - state of the art and general considerations. Herpetologia Sardiniae (Claudia Corti, ed.), Edizioni Belvedere. 2008 (504 S.).
- Pinya, S. & Carretero M.A.: The Balearic herpetofauna: a species update and review on the evidence. Acta Herpetologica 6 (1). 2011, S. 59–80.
- Rato, C., Resen-Des, R., Tristao Da Cunha, R., & James Harris, D.: First record of Tarentola substituta JOGER, 1984, and genetic identification of Tarentola mauritanica (LINNAEUS, 1758), in the Azores. Herpetozoa 27 (3/4). 2015, S. 182–187.
- Mediani, M., Brito, J. C., & Fahd, S.: Atlas of the amphibians and reptiles of northern Morocco: updated distribution and patterns of habitat selection. Basic and Applied Herpetology, 29. 2015, S. 81–107.
- Boettger, O.: Liste der von Hrn. Dr. med. W. Kobelt in Algerien und Tunesien gesammelten Kriechtiere. In W. Kobelt (editor), Reiseerinnerungen aus Algerien und Tunis. Berichte der Senckenbergischen Naturforschungs Gesellschaft 1885. 1885, S. 452–475.
- Sarra, F., Nabil, A., Rached, G., & Khaled, S.: Relationships of the Moorish Gecko Tarentola mauritanica sensu lato (Reptilia, Phyllodactylidae) populations in Tunisia: morphometric and karyological assessment. In Annales Zoologici (Vol. 63, No. 1) Museum and Institute of Zoology, Polish Academy of Sciences. 2013, S. 149–156.
- Geniez, P., Mateo, J.A., Geniez, M. & Pether, J.: The amphibians and reptiles of the Western Sahara (former Spanish Sahara) and adjacent regions. Edition Chimaira, Frankfurt, 228 S. [reviewed in Reptilia GB 44: 81]. 2004.
- Schleich, H.H., Kästle,W. & Kabisch, K.: Amphibians and Reptiles of North Africa. Koeltz, Koenigstein. 1996 (627 S.).
- Buchert, I. & Buchert, P.: Die Gialova-Lagune. elaphe 2011(1), Jahrgang 19. 2011, S. 26–32.
- Mačát, Z., M. Starcová, J. Červenka, D. Jablonski & M. Šandera: A molecular assessment and first record of Tarentola mauritanica (Squamata: Phyllodactylidae) on Corfu, Greece. Salamandra 50 (3). 2004, S. 172–176.
- Mizerakis, V. & Strachinis, I.: New record of Tarentola mauritanica (Squamata: Phyllodactylidae) from Lesvos island, Greece. Herpetology Notes, 10. 2017, S. 157–159.
- Báez, M. & Biscoito, M.: First record of Tarentola mauritanica (Linneus, 1758) from the island of Madeira. In Macaronesian Congress. 1993.
- Jesus, J., Lemos, A., Gonçalves, R. & Brehm, A.: First record of Tarentola mauritanica (Linnaeus, 1758) on Porto Santo Island. Herpetozoa, 20(3/4). 2008, S. 175–177.
- Achaval, F. & Gudynas, E.: Hallazgo de Tarentola mauritanica (L., 1758) (Lacertilia, Gekkonidae) en el Uruguay. Boletín de la Sociedad Zoológica del Uruguay (2a época), 1, 7-10. 1983.
- Baldo, D., Borteiro, C., Brusquetti, F., García, J. E. & Prigioni, C.: Reptilia, Gekkonidae, Hemidactylus mabouia, Tarentola mauritanica: Distribution extension and anthropogenic dispersal. Check List, 4(4). 2008, S. 434–438.
- Huerta-Vera, S.: Registros de Gecko Mediterráneo, Tarentola mauritanica (Linnaeus 1758) (Squamata, Phyllodactylidae), en zona semi-urbana de Peñalolén, Región Metropolitana. Boletín Chileno de Herpetología. 3. 2016, S. 24–25.
- Barts, M.: Unusual day shelter of Spanish Moorish Gecko. Sauria 29 (4). 2007, S. 30.
- Tlili, W., Delaugerre, M., Ouni, R., & Nouira, S.: Distributional review of the genus Tarentola (Reptilia, Sauria) in Tunisia (North Africa). Herpetology Notes, 5. 2012, S. 485–492.
- Pasteur, G. & Girot, B.: Les tarentes de l'Ouest africain. II.-Tarentola mauritanica. Bull. Soc. Sc. nat. phys. Maroc, 40(4). 1960, S. 309–322.
- Geniez, P., Escatllar, J., Crochet, P. A., Mateo, J. A., & Bons, J.: A new form of the genus Tarentola from north-western Africa. Herpetozoa, 12. 1999, S. 187–194.
- Harris, D. J., Batista, V., Carretero, M. A., & Ferrand, N.: Genetic variation in Tarentola mauritanica (Reptilia: Gekkonidae) across the Strait of Gibraltar derived from mitochondrial and nuclear DNA sequences. Amphibia-Reptilia, 25(4). 2004, S. 451–459.
- Rato, C., Carranza, S., Perera, A., Carretero, M. A., & Harris, D. J.: Conflicting patterns of nucleotide diversity between mtDNA and nDNA in the Moorish gecko, Tarentola mauritanica. Molecular Phylogenetics and Evolution, 56(3). 2010, S. 962–971.
- Gil, M.J., Pérez-Mellado, V. & Guerrero, F.: Eine vergleichende Studie des Nahrungserwerbs von Tarentola mauritanica (Reptilia: Gekkonidae) in Habitaten auf dem Festland und auf Inseln. Sauria 15. 1993, S. 9–17.
- Hódar, J. A., Pleguezuelos, J. M., Villafranca, C., & Fernández-Cardenete, J. R.: Foraging mode of the Moorish gecko Tarentola mauritanica in an arid environment: inferences from abiotic setting, prey availability and dietary composition. Journal of arid environments, 65(1). 2006, S. 83–93.
- Werner, Y.L., Bouskila, A., Davies, S.J. & Werner, M.: Observations and comments on active foraging in geckos. Russian Journal of Herpetology 4. 1997, S. 34–39.
- Ulber, T. & E. Ulber: Auf der Mauer, auf der Lauer - Erfahrungen mit der Zimmerhaltung von verschiedenen Geckos. - Sauria, Berlin, 9(2). 1987, S. 7–12.
- Gramentz, D.: Zur Ökologie und Ethologie von Tarentola mauritanica mauritanica (LINNAEUS 1758). Sauria 24 (4). 2002, S. 13–18.
- Gil, M.J., Guerrero, F., Pérez-Mellado, V.: Seasonal variation in diet composition and prey selection in the Mediterranean gecko Tarentola mauritanica. Israel Journal of Zoology 40. 1994, S. 61–74.
- Gil, M.J.: Estudio de la comunidad de saurios de la vertiente meridional de la Sierra de Gredos. Ph.D. Dissertation, Universidad de Salamanca, Salamanca. 1992.
- Cencetti, T., Poli, P., Mele, M., & Zuffi, M. A.: Preliminary results on tail energetics in the Moorish gecko, Tarentola mauritanica. Acta Herpetologica, 6 (1). 2011, S. 101–103.
- Bateman, P.W. & Fleming, P.A.: To cut a long tail short: a review of lizard caudal autotomy studies carried out over the last 20 years. J. Zool., Lond. 277. 2009, S. 1–14.
- Geniez, P., Escatllar, J., Crochet, P. A., Mateo, J. A., & Bons, J.: A new form of the genus Tarentola from north-western Africa. Herpetozoa, 12. 1999, S. 187–194.
- Nettmann, H. K., & Rykena, S.: Verhaltens- und fortpflanzungsbiologische Notizen über kanarische und nordafrikanische Tarentola-Arten. Bonn. Zool. Beitr, 36(3/4). 1985, S. 287–305.
- Nettmann, H.K. & Rykena S.: Mauergeckos (Tarentola mauritancia), die ihre Eier im Sand vergraben (Reptilia: Sauria: Gekkonidae). – Salamandra 15. 1979, S. 53–57.
- Atzori, A., Berti, F., Cencetti, T., Fornasiero, S., Tamburini, M., & Zuffi, M. A.: Advances in methodologies of sexing and marking less dimorphic gekkonid lizards: the study case of the Moorish gecko, Tarentola mauritanica. Amphibia-Reptilia, 28(3). 2007, S. 449–454.
- Kreiner, G.: Schlangen Europas. Edition Chimaira. Frankfurt am Main 2007.
- Machts, T.: Zum Fressen gern! Reptilia Nr. 99, Jahrgang 18(1). 2013, S. 5.
- Tarentola mauritanica in der Roten Liste gefährdeter Arten der IUCN 2019-1. Eingestellt von: Vogrin, M., Corti, C., Perez-Mellado, V., Sá-Sousa, P., Cheylan, M., Pleguezuelos, J., Baha El Din & S. Martínez-Solano, I., 2009. Abgerufen am 11. Juli 2019.
- Zawadzki, M.: Ibiza – Sonne, Strand und Eidechsen. Terraria/Elaphe 2014 (4). 2014, S. 16–21.
- Zawadzki, M.: Mallorca – Die verkannte Insel. Terraria/Elaphe 2014 (4). 2014, S. 34–39.