Katabolitrepression
Die Katabolitrepression ist ein Mechanismus zur Genregulation. Sie dient der Hemmung der Produktion von bestimmten katabolischen Enzymen einer Zelle, so lange wie ein generell energetisch günstigeres Substrat – typischerweise eine andere Kohlenstoffquelle – eines alternativen katabolischen Stoffwechselweges in signifikanter Menge vorhanden ist.[1]
Prinzip
Bei der Katabolitrepression bewirkt nach der ursprünglichen Annahme ein Katabolit des Glucosestoffwechsels eine Hemmung der Genexpression bestimmter, für die Verarbeitung anderer Kohlenstoffverbindungen relevanter Gene, z. B. im Lac-Operon.[2] Jedoch wurde nach der Benennung des beobachteten Effekts entdeckt, dass nicht ein Katabolit der Glucose die Genexpression hemmt, sondern der second messenger cAMP, der bei Glucosemangel gebildet wird und an das Catabolite activator protein (CAP, auch cAMP response protein,[2] CRP) bindet. Nach der cAMP-Bindung wirkt das dimere CAP-Protein als Aktivator der Genexpression.[2] Glucose führt zu einer erniedrigten cAMP-Konzentration.[2] Die Katabolitrepression ist somit eine Aktivierung der Genexpression bei Abwesenheit des Substrats. Nebenbei besitzt das lac-Operon auch den Lac-Repressor, der bei Bindung von Allolactose (einem Stoffwechselprodukt der Lactose) den Promotor freigibt.[2]
Erstmals gezeigt wurde dieser Mechanismus am Beispiel der Glucose, weshalb er früher auch Glucoseeffekt genannt wurde. Aber auch andere Kohlenstoffquellen können für Katabolitrepression verantwortlich sein.
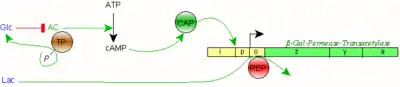
1. cAMP aktiviert bei Glc-Mangel das CAP-Protein, welches direkt an den Promotor (p) bindet und dessen Aktivierung unterstützt;
2. Allolactose bindet an ein Repressorprotein (REP); dieses löst sich daraufhin von der Operator-Sequenz (o) und gibt die Transkription der Geneinheit lacZYA frei.
Bei Verfügbarkeit von Glc sinkt der cAMP-Spiegel durch Inhibition der Adenylylzyklase (AC).
Man bemerkte, dass bestimmte Stoffwechselwege eines Bakteriums gehemmt waren (also die Synthese bestimmter Enzyme ausblieb), wenn Glucose im Medium vorhanden war. Bei Abwesenheit bzw. nach Verbrauch der Glucose wurden die vorher reprimierten Enzyme exprimiert. Besonders deutlich wird dies beim Wachstum von Escherichia coli in einem Medium, welches Lactose und Glucose enthält. Zunächst wächst die Kultur relativ schnell und verbraucht die Glucose, es schließt sich eine kurze Phase ohne Wachstum (lag-Phase oder Latenzphase) an, und anschließend geht das Wachstum unter Lactoseverbrauch weiter, allerdings langsamer. Beim Wachstum auf Lactose wird bei E. coli das Lactose-Operon induziert. Zunächst unterblieb diese Induktion jedoch zugunsten der Verwertung der im Medium vorhandenen Glucose. Die kurze lag-Phase zwischen vollständiger Verwertung der Glucose und Beginn der Lactoseverwertung beschreibt somit den Zeitraum, in dem das Lactose-Operon induziert wurde und die lactoseverwertenden Enzyme gebildet wurden. Ein solches Wachstumsverhalten mit zwei getrennten exponentiellen Phasen wird auch diauxisches Wachstum oder kurz Diauxie genannt.
Der Mechanismus, der diesem Phänomen zugrunde liegt, ist die Katabolitrepression. Glucose hemmt die Adenylatcyclase, ein Enzym, welches aus ATP cyclisches Adenosinmonophosphat (cAMP) synthetisiert. cAMP ist ein universeller intrazellulärer Botenstoff bzw. Signalmolekül (second messenger) und reguliert viele zelluläre Vorgänge. Eine wichtige Funktion von cAMP im hier beschriebenen Kontext ist die Bindung an CAP (Katabolit-Aktivatorprotein) genannt. Bei katabolitreprimierten Genen kann die RNA-Polymerase nur dann an die DNA binden und die Transkription einleiten, wenn vorher an die DNA CAP gebunden wurde. Diese Anlagerung kann nur bei einer vorherigen Bindung an cAMP stattfinden. Bei einem Absinken der intrazellulären cAMP-Konzentration können CAP-abhängige Gene nicht transkribiert werden und deren Produkte nicht gebildet werden. Das Lactose-Operon unterliegt ebendiesem Kontrollmechanismus, es ist aber nicht das einzige. Da einerseits viele Operons mittels des cAMP-Levels kontrolliert werden, andererseits Operons in der Regel eigene Regulatoren besitzen (im Falle des lac-Operons: Allolactose und der Lac-Repressor), ermöglicht dieses einfache Regulationssystem eine Modulation des Zellstoffwechsels.
Weblinks
- Jennifer McDowall/Interpro: Protein Of The Month: Catabolite Activator Protein. (englisch)
Einzelnachweise
- Katharina Munk, Petra Dersch: Mikrobiologie : 43 Tabellen. Thieme, Stuttgart [u. a.] 2008, ISBN 3-13-144861-X, S. 271.
- Jeremy M. Berg, John L. Tymoczko, Lubert Stryer: Biochemistry. 5. Auflage. Freeman, New York 2002, ISBN 0-7167-4684-0, Kapitel 31.1.6. Online verfügbar beim NCBI Bookshelf.