Metathalamus
Als Metathalamus (gr. μετά meta 'über, nach, hinter'; 'Nachthalamus') wird ein Teil des Gehirns der Säugetiere, genauer des Zwischenhirns (Diencephalon) bezeichnet, nämlich ein Bereich des Thalamus an dessen dorsalem (hinterem) Abschnitt. Der Metathalamus besteht aus den beiden, je paarigen, seitlichen und mittleren Kniehöckern (Corpora geniculata laterale et mediale).
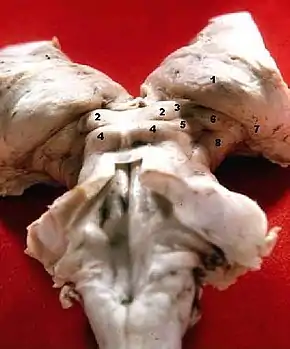
Corpus geniculatum laterale (7)
Corpus geniculatum laterale
Der seitliche Kniehöcker (Corpus geniculatum laterale, CGL) ist ein Kerngebiet, in dem etwa 90 % der Axone der Sehbahn (Tractus opticus) enden. Die Neurone des CGL erhalten jedoch nicht nur Input von der Netzhaut, sondern auch von anderen Thalamuskernen und von Neuronen des CGL selbst, sowie von der Großhirnrinde und den oberen Hügeln des Mittelhirns. All diese Informationen werden hier integriert, modifiziert und zu einem großen Teil über die Sehstrahlung (Radiatio optica) an den visuellen Cortex weitergegeben. Daneben bestehen Efferenzen zu Kerngebieten im Colliculus superior, zu den Nuclei pretectales und zum Nucleus suprachiasmaticus des Hypothalamus.
(Die übrigen 10 % der Sehnervenfasern enden direkt an Kernen in den Colliculi superiores, in der Area praetectalis oder im Hypothalamus. Ihre Informationen dienen unter anderem den reflektorischen Kopf- und Augenbewegungen, dem Pupillenreflex und dem Tag-Nacht-Rhythmus.)
Die Kerngebiete des Corpus geniculatum laterale zeigen jederseits ein Aufbau aus sechs kappenartig gekrümmten, in etwa konzentrisch angeordneten Schichten, für die sich ein großzelliges (magnozelluläres) Gebiet (Nucleus ventralis corporis geniculati lateralis) und ein kleinzelliges (parvozelluläres) Gebiet (Nucleus dorsalis corporis geniculati lateralis) unterscheiden lassen.
Aufteilung in Schichten
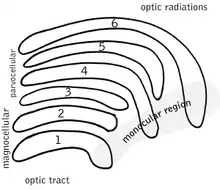
Die Schichten 1, 4 und 6 sind Ziel kontralateraler Projektionen über den Tractus opticus.
In den beiden Schichten 1 und 2 liegen größere Zellen, nach dorsal folgen parvozellulär Schicht 3 bis 6.
(Koniozelluläre Unterschichten, jeweils ventral anliegend, nicht dargestellt.)
Zuordnung der Gesichtsfeldhälfte
Das CGL ist aus sechs Schichten aufgebaut, welche von ventral nach dorsal mit Schicht 1 bis 6 bezeichnet werden. Die Schichten 1, 4 und 6 erhalten Input vom kontralateralen Auge, die Schichten 2, 3 und 5 vom ipsilateralen Auge, und zwar jeweils über die dem CGL gegenüberliegende Hälfte des Gesichtsfeldes. Das rechte CGL erreichen also Informationen über die linke Gesichtsfeldhälfte, die aus dem linken Auge stammen (Schichten 1, 4 und 6), und Informationen über die linke Gesichtsfeldhälfte aus dem rechten Auge (Schichten 2, 3 und 5). Somit erhält jede Gehirnhälfte wohl Informationen von beiden Augen, doch vorerst nur über die eine gegenseitige Gesichtsfeldhälfte (siehe auch: Kontralateralität des Vorderhirns). Diese Trennung bleibt in den Projektionen des CGL zum primären visuellen Cortex (V1, Area striata, Area 17) erhalten und erreicht so die neokortikale Schicht 4 des visuellen Cortex. Erst in der weiteren kortikalen Verarbeitung werden die Informationen über beide Gesichtsfeldhälften dann in unterschiedlich starkem Ausmaß zusammen verarbeitet.
Die beiden ventralen Schichten 1 und 2 des CGL werden als magnozelluläre Schichten (oder auch ventraler Kern), die Schichten 3 bis 6 als parvozelluläre Schichten (oder auch dorsaler Kern) bezeichnet. Sie erhalten Input von unterschiedlichen Typen retinaler Ganglienzellen. Zwischen den sechs Schichten befinden sich die koniozellulären Schichten sehr kleiner Zellen, deren Funktion noch nicht vollständig geklärt ist. Sie erhalten Input von retinalen Ganglienzellen des bistratified-Typs.
Magnozelluläre Schichten (Nucleus ventralis CGL)
Die Neurone der beiden ventralen Schichten des CGL zeichnen sich durch größeren Zellkörper, umfangreichere Dendritenbäume und stärkere Axone im Vergleich zu den Neuronen der anderen Schichten aus. Der größere Durchmesser ihrer Axone bewirkt eine sehr hohe Leitungsgeschwindigkeit zu nachgeschalteten Einheiten. Die Inputzellen dieser Schicht – die magnozellulären retinalen Ganglienzellen vom parasol-Typ – zeichnen sich durch ein großes rezeptives Feld aus. Sie sind jedoch gänzlich insensitiv für Farbwahrnehmung, da ihre rezeptiven Felder sowohl im Zentrum als auch der Peripherie aus derselben Zapfenart bestehen (Farbwahrnehmung ist nur möglich, wenn Zentrum und Peripherie eines rezeptiven Feldes Input aus verschiedenen Zapfenarten erhalten). Die Funktion des magnozellulären Systems hat man insbesondere mit Läsionsstudien an Affen untersucht: Dabei wurde die magno- bzw. parvozellulären Schichten selektiv durch pharmazeutische Methoden deaktiviert. Affen, bei denen die magnozellulären Schichten des CGL ausgeschaltet waren, zeigten deutliche Defizite bei der Bewegungs-, Orts- und Geschwindigkeitswahrnehmung. Farbsehen und visuelle Genauigkeit waren jedoch nicht beeinträchtigt. Daher sieht man in den magnozellulären Schichten einen wesentlichen Input zur „dorsalen Route“ der visuellen Verarbeitung im Gehirn, welche v. a. Bewegungs-, Ort- und Handlungswahrnehmung verarbeitet. Die dorsale Route umfasst ausgehend von V1 verschiedene Bereiche in Richtung Parietalcortex, wie Area MT, MST, MIT.
Parvozelluläre Schichten (Nucleus dorsalis CGL)
In der dritten bis sechsten Schicht des CGL befinden sich parvozelluläre Neurone mit kleineren Zellkörpern, eingeschränkteren Dendritenbäumen und dünneren Axonen (bedingt langsamere Leitungsgeschwindigkeit). Sie erhalten Input von parvozellulären retinalen Ganglienzellen des midget-Typs, welche ein kleineres rezeptives Feld aufweisen und damit eine höhere visuelle Auflösung ermöglichen. Farbwahrnehmung ist nur mit dem parvozellulären System möglich. Das Zentrum eines rezeptiven Feldes einer parvozellulären Ganglienzelle besteht aus einer anderen Zapfenart als ihre Peripherie. Über den Vergleich der Erregungsstärke der Zapfenarten kann eine Farbe wahrgenommen werden. Es existieren drei Zapfenarten, welche jeweils für kurz-, mittel- oder aber langwelliges Licht (blau-, grün- oder rot) die größte Empfindlichkeit zeigen.
Das Parvozelluläre System ist für eine Wahrnehmung mit visueller Auflösung feinerer Akkuratesse (Größe, Form, Textur) nötig, ebenso für die Farbwahrnehmung. Es gilt als wichtigster Input für die „ventrale Route“ der visuellen Verarbeitung im Gehirn, welche ausgehend von V1 Richtung ventral zum inferioren Temporallappen verläuft (z. B. V4, Gyrus fusiformis). Es ist zuständig für Form-, Schärfe-, Farbwahrnehmung und Objekterkennung.
Corpus geniculatum mediale
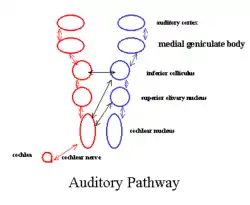
Der mediale Kniehöcker (Corpus geniculatum mediale) ist Umschaltstation der Hörbahn. Deren Informationen gelangen über den Lemniscus lateralis und Colliculus inferior zum Kniehöcker und von dort über die Radiatio acustica zum auditiven Cortex im Temporallappen. Das Corpus geniculatum mediale gliedert sich anatomisch und funktionell in drei Teile.
Pars ventralis
Die Pars ventralis ist der Hauptteil des auditorischen Thalamus. Sie umfasst die gesamte spezifische (lemniskale) Komponente der Hörbahn, also den Hauptstrang der akustischen Informationsverarbeitung. Sie erhält aufsteigende Fasern vom zentralen Kern des gleichseitigen Colliculus inferior und zieht mit ausgehenden Fasern zum gleichseitigen primären auditorischen Cortex (AI) und vorderen (anterioren) auditorischen Feld (AAF). Im Zuge einer Rückkopplung erhält sie absteigende Fasern von allen am Hörsystem beteiligten Arealen des Cortex.
Die Neurone und der Einzugsbereich ihrer Dendriten sind in Schichten (Laminae) geordnet. Mehrere benachbarte Neuronen-Schichten bilden einen funktionellen Schichtenverbund. In der Fläche eines Verbunds besteht eine Fein-Skalierung nach akustischen Frequenzen, während von Verbund zu Verbund eine Grob-Skalierung im Abstand einer Oktave vorliegt.[1]
Die Funktion der Pars ventralis besteht in der Kombination jeder Art von frequenzspezifischer Information und ihrer Modulation durch absteigenden Input vom Kortex, bevor diese zur Weiterverarbeitung an den Kortex geliefert wird. Ein spezieller Effekt von Anatomie und Funktion ist möglicherweise die in der Musik bekannte Oktavidentität.
Pars dorsalis
Die Pars dorsalis ist Teil der unspezifischen (extralemniskalen) Komponente der Hörbahn, die auditive und nicht-auditive Information kombiniert. Sie erhält aufsteigende Fasern vom dorsalen Kortex des gleichseitigen Colliculus inferior und vom somatosensorischen System. Sie zieht mit ausgehenden Fasern zum gleichseitigen sekundären und tertiären auditorischen Cortex (AII u. a.). Im Zuge einer Rückkopplung erhält sie absteigende Fasern von allen am Hörsystem beteiligten Arealen des Cortex.
Die Funktion der Pars dorsalis besteht in der Kombination jeder Art von nicht-frequenzspezifischer auditiver mit nicht-auditiver Information. Auch hier unterliegt dieser Prozess einer Modulation durch absteigenden Input vom Kortex.
Pars medialis
Die Pars medialis ist ebenfalls Teil der unspezifischen (extralemniskalen) Komponente der Hörbahn. Sie erhält aufsteigende Fasern von allen drei Hauptabteilungen des gleichseitigen Colliculus inferior, von der oberen Olive (Nucleus olivaris superior), vom Colliculus superior und vom Gleichgewichtssystem. Sie zieht mit ausgehenden Fasern zu allen Teilbereichen des auditorischen Cortex (AI, AII u. a.) und auch zur Amygdala, einen Kerngebiet des Limbischen Systems. Im Zuge einer Rückkopplung erhält sie absteigende Fasern von allen am Hörsystem beteiligten Arealen des Cortex.
Die Funktion der Pars medialis besteht in der Kombination jeder Art von nicht-frequenzspezifischer auditiver mit nicht-auditiver Information. Durch die Verbindung mit der Amygdala ist dieses Kerngebiet unter anderem beteiligt an akustischer Konditionierung von Gefühlen. Ferner unterliegt es, wie die anderen Hauptteile des Corpus geniculatum mediale, einer Modulation durch absteigenden Input vom Kortex.[2]
Einzelnachweise
- J. S. Cetas, R. O. Price, D. S. Velenovsky, D. G. Sinex, N. T. McMullen: Frequency organization and cellular lamination in the medial geniculate body of the rabbit. In: Hearing research. Band 155, Nummer 1–2, Mai 2001, ISSN 0378-5955, S. 113–123, PMID 11335081.
- James O. Pickles: Auditory pathways: anatomy and physiology. In: Gastone G. Celesia, Gregory Hickok (Hrsg.): The Human Auditory System: Fundamental Organization and Clinical Disorders. Band 129 von Handbook of Clinical Neurology, Burlington: Elsevier Science 2015, 722 S., S. 3–25, ISBN 0444626298, S. 13–15.