Doushantuo-Formation
Die Doushantuo-Formation (chinesisch 陡山沱组) ist eine aus Sedimenten des Ediacariums entstandene Formation in den chinesischen Provinzen westliches Hubei und nordöstliches Guizhou (Kreis Weng’an). Sie ist bekannt für ihre sehr alten und vorzüglich erhaltenen Fossilien mit sehr frühen Metazoa und eventuellen Bilateria.
Geologie
Die Geologie im Ablagerungsbereich der Doushantuo-Formation war bereits im Jahr 1924 von J. S. Lee und Y. T. Chao[1] erstmals beschrieben worden. Die Formation bildet Teil der Sedimente des endneoproterozoischen Siniums (oder sinischen Systems), die zusammen mit dem Prä-Sinium dem metamorphen archaischen und paläoproterozoischen Grundgebirge des Jangtse-Kratons diskordant auflagern und mit 10 bis 20° nach Südost einfallen. Die Sedimente entstanden als Antwort auf das Auseinanderbrechen Rodinias, das im Zeitraum 830 bis 750 Millionen Jahre erfolgt war (so hatte sich der Jangtse-Kraton damals von Australien und/oder Indien getrennt). Im Cryogenium wurden dann im Zeitraum 750 bis 635 Millionen Jahre glaziale und interglaziale Sedimente abgesetzt, die den Übergang vom anfänglichen Rift- (Nanhua-Rift) zum Driftstadium dokumentieren.
Im Bereich der Schluchten des Jangtsekiang nordwestlich von Yichang bildet das Grundgebirge eine nach Nordnordost ausgelängte, rund 60 Kilometer lange und 30 Kilometer breite domartige Aufwölbung (Huangling-Antiklinal), die zum überwiegenden Teil vom neoproterozoischen Huangling-Granit ausgefüllt wird. Der Granit war vor 819 ± 7 Millionen Jahren in das Grundgebirge eingedrungen.
Die neoproterozoische Sedimentabfolge des passiven Kontinentalrandes des Jangtse-Kratons beginnt mit der 400 Meter mächtigen Liantuo-Formation, die aus rund 750 Millionen Jahre alten Sandsteinen des Cryogeniums aufgebaut wird.[2] Darüber legen sich rund 100 Meter an grünen Tilliten der Nantuo-Formation, die im Mittelabschnitt eine rote Sandsteinlage enthält.
Stratigraphie
Die Doushantuo-Formation überlagert die Tillite der Nantuo-Formation, die am Ende der Marinoischen Vereisung abgesetzt worden waren. Diese das Cryogenium beendenden Geschiebemergel (Diamiktite) konnten mit zirka 654 bis 635 Millionen Jahren datiert werden. Die Doushantuo-Formation wird ihrerseits von Schwarzschiefern sowie massiven Dolomiten und Kalken der Dengying-Formation überdeckt, welche aus dem ausgehenden Ediacarium stammen.[3]
Die sehr stark kondensierte Doushantuo-Formation weist in ihrem Verbreitungsgebiet große Mächtigkeitsschwankungen (zwischen 40 und 300 Meter, durchschnittlich 160 bis 230 Meter) und auch bedeutende Fazieswechsel auf. An ihrer Typlokalität in den Schluchten des Jangtse bei Jiulongwan erreicht sie über 200 Meter an Mächtigkeit und kann in vier Member unterteilt werden:
Das nur 2 bis 6 Meter mächtige Member I besteht aus dolomitischen Hutkarbonaten (Englisch cap carbonates), die durch zahlreiche ungewöhnliche Sediment- und Diagenesestrukturen wie mit Quarz verfüllte horizontale Schrumpfungsrisse (Englisch sheet cracks), Makropeloide, Teepeestrukturen und Barytfächer gekennzeichnet werden. In ihm lassen sich drei Horizonte erkennen, ein gestörter, ein lagiger und ein siltiger Kalk-Dolomitverband. Das rund 80 bis 150 Meter mächtige Member II ist eine Wechselfolge aus dünnbankigen Dolomikriten, kohlenstoffreichen tonigen Dolomiten (mit reichlichen Kieselknollen) und kohlenstoffreichen schwarzen, kalkhaltigen Schiefertonen. In den enthaltenen erbsengroßen Phosphorit-Chert-Knollen, die frühdiagenetischen Ursprungs sind und vorwiegend in der unteren Hälfte auftreten, wurden ausgiebig Acritarchen und andere Fossilien gefunden. Member III wird 60 bis 80 Meter mächtig und besteht aus massivem und mittel- bis dickbankigem, kieseligen Dolomit mit dünnen, an Mikrofossilien reichen Chertbänden. Überlagert wird der Dolomit von einer Wechselfolge aus dünn- bis dickbankigen tonigen Kalken und mittel- bis dickbankigen dolomitischen Tonsteinen; Intraklasten und Karbonatkonkretionen können vorhanden sein. Das Member IV ist ein 5 bis 10, manchmal bis 20 Meter mächtiger, mit organischem Kohlenstoff angereicherter Schwarzschiefer (kalkiger bis kieseliger Schieferton oder Tonstein), der den Schiefertonen des Member II sehr ähnlich ist, jedoch keinerlei Sedimentstrukturen aufweist; vorhandene Karbonatkonkretionen erreichen den Dezimeterbereich; zugegen sind ferner Pyrit und Baryt.
Eine ausführliche Beschreibung der Lithologie findet sich bei McFadden und Kollegen (2008).[4]
Sequenzstratigraphisch kann die Doushantuo-Formation in zwei Vollzyklen (Transgression-Regression) und die Hälfte eines dritten Zyklus unterteilt werden, wobei die Grenze zwischen Sequenz 1 und Sequenz 2 in der Mitte von Member II verläuft. Die Grenze zwischen Sequenz 2 und der unvollständigen Sequenz 3 liegt innerhalb von Member III am Umschwung von dolomitischer zu kalkiger Sedimentation.[5] Ferner lassen sich oft zwei Flutungsflächen (Englisch flooding surfaces) beobachten – die untere ist mit der Grenze von Member II zu Member III identisch, die obere liegt etwas oberhalb der Grenze von Member III zu Member IV.
Ablagerungsbedingungen
Die insgesamt regressive Doushantuo-Formation war auf einem in einen tieferen Beckenbereich überleitenden Karbonatschelf im Süden und Südosten des Jangtse-Kratons abgelagert worden.[4] Member II war unter ruhigen Bedingungen unterhalb der Sturmwellenbasis sedimentiert worden, Member III zeigt jedoch wesentlich bewegtere Strukturen und lässt auf subtidale Bedingungen schließen.
Der Schelfrand umgab tiefergelegene Lagunen, die sich vom Gezeitenbereich bis zum offenen Meer erstreckten. Die Wassermassen in den Lagunen nahmen unter Anreicherung von Schwefelwasserstoff hin und wieder einen anoxischen bzw. euxinischen Charakter an. Isotopen- und Elementkonzentrationen in den Lagunensedimenten geben als Proxy diese Schwankungen zu erkennen, welche wahrscheinlich zur speziellen Fossilerhaltung in der Doushantuo-Formation beitrugen.
Die Schelfmorphologie war bereits während des Nanhua-Riftsystems angelegt worden. Sie erklärt die Variabilität der Doushantuo-Formation. Während der folgenden Dengying-Formation entwickelte sich die Jangtse-Plattform schließlich im Zeitraum 551 bis 542 Millionen Jahre zu einem in ein Tiefwasserbecken (Liuchapo-Formation) übergehenden offenen Schelf.
Die innerhalb der Formation so häufig auftretenden Chertknollen sind frühdiagenetische Bildungen, die an der Grenzfläche Sediment/Meerwasser ihren Ursprung nahmen.
Geochemie
Kohlenstoffisotopen
In der Doushantuo-Formation lassen sich vier negative δ13C-Anomalien erkennen: CANCE an der Basis, WANCE an der Grenze von Sequenz 1 zu Sequenz 2, BAINCE innerhalb von Sequenz 2 und DOUNCE an der Grenze von Sequenz 2 zu Sequenz 3 im Hangenden.[5]
Im Hutdolomit des Member I liegt CANCE mit negativen δ13C-Werten, wie sie für das frühe Ediacarium typisch sind (generell bis - 5 ‰). Die Anomalie WANCE ist relativ schwach ausgeprägt und erreicht - 2 ‰. Im oberen Abschnitt des Member III tritt mit BAINCE erneut eine negative δ13C-Anomalie auf, mit Werten bis zu - 8 ‰. Die letzte Anomalie DOUNCE ist mit - 10 bis - 13 ‰ sehr kräftig und wird mit der Shuram- bzw. der Wonoka δ13C-Anomalie korreliert. Zum Top des Member IV steigen die δ13C-Werte dann wieder gegen 0 an. Geochemische Analysen lassen für DOUNCE im Member IV auf ein bedeutendes Sauerstoffanreicherungsereignis im Weltmeer schließen.
Diese letzte Anomalie ist globaler Natur und tritt weltweit auf, sie kann aber mit keinem größeren bekannten Ereignis wie beispielsweise einem Massensterben in Verbindung gebracht werden. Möglicherweise handelt es sich hierbei um eine Rückkoppelung zwischen evolutiver Innovation und der chemischen Zusammensetzung des Meerwassers. So könnten die neu aufgetretenen Vielzeller (Metazoa) den für ihr Wachstum benötigten Kohlenstoff dem Wasser entzogen haben, wodurch wiederum die Sauerstoffkonzentration auf indirekte Weise erhöht wurde. Die gestiegenen Sauerstoffwerte ermöglichten nun ihrerseits weitere Evolutionsschritte bei den Metazoa.[6]
Sauerstoffisotopen
Das Sauerstoffisotopenverhältnis δ18O zeigt bis auf einige kleinere Ausnahmen eine schwach negative Korrelation mit dem δ13C-Verlauf. Deutlich ausgeprägt sind die Anomalien CANCE und DOUNCE mit - 15 bzw. - 8 ‰. Der restliche Kurvenverlauf bewegt sich meist zwischen - 5 und 0 ‰. Im Grenzbereich zwischen Member II und Member III werden positive δ18O-Werte erreicht. Dies deutet auf eine klare Abkühlung des Klimas und markiert wahrscheinlich die Gaskiers-Eiszeit.[7]
Strontiumisotopen
Die Strontiumanomalien sind im Hutdolomit sehr deutlich ausgeprägt. So sinkt das radiogene 87Sr/86Sr-Verhältnis von 0,712 an der Basis drastisch auf 0,708 ‰ ab. Letzterer Wert wird dann unter leichten Schwankungen im Rest der Formation beibehalten. Bezeichnend sind auch die δ88Sr-Werte, die von sehr leichtem - 0,42 auf + 0,38 ‰ ansteigen. Dies lässt sich wie folgt erklären: nach dem Ende der Marinoischen Eiszeit hatten sich extreme Treibhausbedingungen eingestellt und die damit verbundene Erwärmung des Weltklimas führte zu einem massiven Abschmelzen der globalen marinoischen Eismassen. Dieses kontinentale Schmelzwasser hatte eine hohe 87Sr/86Sr-Signatur, jedoch einen sehr niedrigen δ88Sr-Wert. Ein Anstieg von Kohlendioxid, bedingt durch vulkanische Ausgasungen sowie eine mögliche Lösung von Methanhydrat bewirkten eine Ausfällung von Karbonaten mit anfangs sehr hohem 87Sr/86Sr und niedrigem δ88Sr. Der darauf folgende Übergang im Hangenden des Hutdolomits zu normalen Strontium-Werten beruhte auf einströmenden Tiefenwasser, dass sich zusehends mit dem Oberflächenwasser vermischte (Tiefenwasser hat normale δ88Sr- und niedrige 87Sr/86Sr-Werte).
Alter
Condon und Kollegen (2005) datierten die Doushantuo-Formation radiometrisch auf das Zeitintervall 635 bis 551 Millionen Jahre.[6] Dies ergibt für die Formation eine relativ niedrige Sedimentationsrate von 3,45 Meter/Million Jahre. Das Hutkarbonat ergab 621 ± 7 Millionen Jahre. Zhang und Kollegen (2005) hatten für das Top der Doushantuo-Formation 555,2 ± 6,1 Millionen Jahre ermittelt.[8] Eine Aschenlage 5 Meter oberhalb des Hutdolomits ergab 632,5 ± 0,5 Millionen Jahre.
Anmerkung: Condon und Kollegen hatten das basale Hutkarbonat mit 635,2 ± 0,6 Millionen Jahren sowie das Hangende des Miaohe-Members mit 551 ± 0,7 Millionen Jahren bestimmt. Mittlerweile hat sich herausgestellt, dass das Miaohe-Member nicht wie angenommen zur Doushantuo-Formation gehört, sondern vielmehr als Liegendes der Dengying-Formation anzusehen ist. An und Kollegen (2015) gehen daher jetzt davon aus, dass die Doushantuo-Formation älter als 560 Millionen Jahre sein muss.[9]
Fossilführung
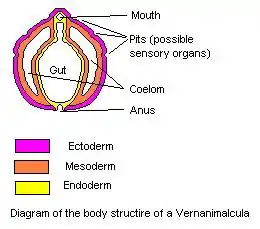
Die Fossilien der Doushantuo-Formation erscheinen in drei taphonomischen Fenstern:
- als kohlige Abdrücke (Englisch carbonaceous compressions) in Schwarzschiefern
- phosphatisiert in Phosphoriten
- verkieselt (silizifiziert) in Cherts.
In Jiulongwan wurden kohlenstoffreiche Abdrücke, die Chuaria ähneln, sowie ein Algenthallus in Schwarzschiefern des Members II entdeckt.[10] In Miaohe treten reichlich kohlige Abdrücke im Schwarzschiefer des Members IV auf.[11] Verkieselte Mikrofossilien sind in kieseligen Knollen oder Linsen sowohl in Member II als auch in Member III anzutreffen.[12]
Die mikroskopisch kleinen Fossilien der Doushantuo-Formation sind durchweg marinen Ursprungs, ihr Erhaltungsgrad ist sehr detailgetreu. Die Struktur der Organismen kann daher selbst noch auf Zellebene untersucht werden. Hieraus erwuchsen neue Einsichten in die Embryo- und Larvenstadien vieler Lebewesen.
Unter den Fossilien finden sich vielzellige Algen[12] und Thallophyten (Seetange), acanthomorphe Acritarchen,[13] Wimpertierchen (Ciliaten),[14] verkieselte Cyanobakterien (als Coccoide und Fäden),[15] verkieselte röhrenförmige Mikrofossilien[16] sowie ausgereifte Schwämme und Nesseltiere (Cnidaria). Bei letzteren handelt es sich möglicherweise auch um Frühformen tabulater Korallen (Tetrakorallen). Daneben treten auch das Taxon Parapandorina – vermutlich der Embryo eines Bilateriums – sowie Eier (Gattung Megasphaera) auf. Manche der vermeintlichen Tierembryos befinden sich im frühen Stadium der Zellteilung und wurden daher oft als Sporen oder Algenzellen interpretiert.[17] Die Eier und Embryos dürften aber vielmehr zu Schwämmen oder Nesseltieren gehören, insbesondere deren ausgereiften Wachstumsformen. Umstritten ist auch nach wie vor das Taxon Vernanimalcula, das als ältester Vertreter der Bilateria vorgeschlagen wurde, von Bengtson und Budd (2004) jedoch als taphonomisches Artefakt angesehen wird.[18]
Es besteht laut Bailey und Kollegen (2007) durchaus die Möglichkeit, dass die angeblichen Embryos und Eier in Wirklichkeit riesige, dem Taxon Thiomargarita ähnelnde Schwefelbakterien sind. Thiomargarita kann bereits ohne optische Hilfsmittel erkannt werden.[19] Diese Sichtweise würde auch die phosphatische Fossilisation erklären, denn in modernen Environments konnte beobachtet werden, dass unter Mitwirkung von Bakterien Phosphat abgeschieden wird. Dem entgegenzuhalten ist, dass neuere Untersuchungen nur wenig Übereinstimmung zwischen den Doushantuo-Fossilien und verwesenden Thiomargarita-Bakterienzellen oder Seeigeleiern feststellen. Problematisch sind auch die dunklen Flecken in den Fossilien, sie als fossile Zellkerne zu interpretieren dürfte jedoch unwahrscheinlich sein.[20]
Folgende Taxa sind in der Doushantuo-Formation gegenwärtig:[21]
- Cyanobakterien - Coccoide:
- Archaeophycus venustus und Myxococcoides
- Cyanobakterien - Filamente:
- Aggregatosphaera miaoheensis, Anomalophyton zhangzhongyingi, Baculiphyca taeniata, Beltanelliformis brunsae, Calyptrina striata, Cucullus fraudulentus, Doushantuophyton lineare, D. quyuani, Enteromorphites siniansis, Glomulus filamentum, Jiuqunaoella simplicis, Konglingiphyton erecta, Liulingjitaenia alloplecta, Longifuniculum dissolutum, Miaohephyton bifurcatum, Obruchevella parva, Oscillatoriopsis amadeus, O. longa, O. obtusa, Polytrichoides lineatus, Protoconites minor, Salome hubeiensis, Sinocylindra yunnanensis, Sinospongia chenjunyuani, S. typica, Siphonophycus capitaneum, S. kestron, S. robustum, S. sinensis, S. solidum und S. typicum[11]
- Vielzellige Algen - Rotalgenthalli:
- Gremiphyca corymbiata, Paramecia incognata, Sarcinophycus papilliformis, S. radiatus, Thallophyca corrugata, T. ramosa, Thallophycoides phloeatus, Wengenia exquisita, W. globosa und W. minuta
- Acanthomorphe Acritarchen:
- Apodastoides basileus, Appendisphaera tenuis, Asterocapsoides densus, A. sinensis, Bacatisphaera baokangensis, Briareus borealis, Carionopora ordinata, Castaneasphaera speciosa, Cavaspina acuminata, C. basiconica, Cymatiosphaeroides kullingii, C. yinii, Dicrospinasphaera zhangii, Distosphaera speciosa, Echinosphaeridium maximum, Eotylotopalla dactylos, E. delicata, Ericiasphaera magna, E. rigida, E. sparsa, E. spjeldnaesii, Filisphaeridium, Goniosphaeridium acuminatum, G. conoideum, G. cratum, Hocosphaeridium scaberfacium, Knollisphaeridium maximum, Leiosphaeridia crassa, L. jacutica, L. tenuissima, Lophosphaeridium, Mastosphaera changyangensis, Meghystrichosphaeridium chadianensis, M. densum, M. gracilentum, M. magnificum, M. perfectum, M. reticulatum, Micrhystridium, Papillomembrana compta, Paratetraphycus giganteus, Polygonium cratum, Polytrichoides, Pustulisphaera membranacea, Sinosphaera rupina, Tanarium anozos, T. conoideum, Tianzhushania ornata, T. spinosa, T. tubifera, Trachyhystrichosphaera, Vulcanisphaera phacelosa und Yangtzitubus semiteres
- Embryos:
- Caveasphaera costata, Megaclonophycus onustus, Megasphaera inornata, M. ornata, Parapandorina raphospissa und Spiralicellula bulbifera.[22]
- Bilateria:
- röhrenförmige Cnidaria:
- Sinocyclocyclicus guizhouensis.
Einzelnachweise
- Zhao Yazeng 赵亚曾 (1898-1929), chinesischer Paläontologe und Geologe.
- Ma, G., Li, H. und Zhang, Z.: An investigation of the age limits of the Sinian System in South China. In: Bulletin of the Yichang Institute of Geology and Mineral Resources (Chinesisch mit englischer Zusammenfassung). Band 8, 1984, S. 1–29.
- Jiang, G., Shi, X., Zhang, S., Wang, Y. und Xiao, S.: Stratigraphy and paleogeography of the Ediacaran Doushantuo Formation (ca. 635-551 Ma) in South China. In: Gondwana Research. Band 19 (4), 2011, S. 831–849, doi:10.1016/j.gr.2011.01.006.
- McFadden, K. A. u. a.: Pulsed oxygenation and biological evolution in the Ediacaran Doushantuo Formation. In: Proceedings of the National Academy of Sciences, USA. Band 105, 2008, S. 3197–3202.
- Zhu, M. u. a.: Carbon isotope chemostratigraphy and sedimentary facies evolution of the Ediacaran Doushantuo Formation in Western Hubei, South China. In: Precambrian Research. Band 225, 2013, S. 7–28, doi:10.1016/j.precamres.2011.07.019.
- Condon, D., Zhu, M., Bowring, S., Wang, W., Yang, A. und Jin, Y.: U-Pb Ages from the Neoproterozoic Doushantuo Formation, China. In: Science. Band 308 (5718), 2005, S. 95–98, doi:10.1126/science.110776.
- Y. Sawaki u. a.: The Ediacaran radiogenic Sr isotope excursion in the Doushantuo Formation in the three Gorges area, South China. In: Precambrian Research. Band 176, 2010, S. 46–64, doi:10.1016/j.precamres.2009.10.006.
- Zhang, S. u. a.: U-Pb sensitive high-resolution ion microprobe ages from the Doushantuo Formation in South China: constraints on late Neoproterozoic glaciations. In: Geology. Band 30, 2005, S. 473–476.
- Zhihui An u. a.: Stratigraphic position of the Ediacaran Miaohe biota and its constraints on the age of the upper Doushantuo δ 13C anomaly in the Yangtze gorges area, South China. In: Precambrian Research. Band 271, 2015, S. 243–253, doi:10.1016/j.precamres.2015.10.007.
- Tang, F., C. Yin, Y. Liu, Z. Wang und L. Gao: Discovery of macroscopic carbonaceous compression fossils from the Doushantuo Formation in eastern Yangtze Gorges. In: Chinese Science Bulletin. Band 50, 2006, S. 2632–2637.
- Xiao, S., X. Yuan, M. Steiner und A. H. Knoll. 2002: Macroscopic carbonaceous compressions in a terminal Proterozoic shale: A systematic reassessment of the Miaohe biota, South China. In: Journal of Paleontology. Band 76, 2002, S. 347–376.
- Xiao, S.: New multicellular algal fossils and acritarchs in Doushantuo chert nodules (Neoproterozoic, Yangtze Gorges, South China). In: Journal of Paleontology. Band 78, 2004, S. 393–401.
- Zhou, C., G. Xie, K. McFadden, S. Xiao und X. Yuan: The diversification and extinction of Doushantuo–Pertatataka acritarchs in South China: Causes and biostratigraphic significance. In: Geological Journal. Band 42, 2007, S. 229–262.
- C.-W. Li, J.-Y. Chen, J. H. Lipps, F. Gao, H.-M. Chi und H.-J. Wu: Ciliated protozoans from the Precambrian Doushantuo Formation, Wengan, South China. In: Vickers-Rich, Patricia und Komarower, Patricia, The Rise and Fall of the Ediacaran Biota (Hrsg.): Geological Society, Special publications. Band 286. London 2007, S. 151–156, doi:10.1144/SP286.11.
- Zhang, Y., L. Yin, S. Xiao und A. Knoll: Permineralized fossils from the terminal Proterozoic Doushantuo Formation, South China. In: Journal of Paleontology. 72 (supplement), 1998, S. 1–52.
- Liu, Pengju u. a.: Silicified tubular microfossils from the Upper Doushantuo Formation (Ediacaran) in the Yangtze Gorges area, South China. In: Journal of Paleontology. Band 83(4), 2009, S. 630–633.
- Yin, L., M. Zhu, A. H. Knoll, X. Yuan, J. Zhang und J. Hu: Doushantuo embryos preserved inside diapause egg cysts. In: Nature. Band 446, 2007, S. 661–663.
- Stefan Bengtson and Graham Budd: Comment on "Small Bilaterian Fossils from 40 to 55 Million Years Before the Cambrian". In: Science. Band 306 (5700), 2004, S. 1291a.
- Bailey, Jake V., Joye, S. B, Kalanetra, K. M., Flood, B. E. und Corsetti, F. A.: Evidence of giant sulphur bacteria in Neoproterozoic phosphorites. In: Nature. Band 445 (7124), 2007, S. 198–201, doi:10.1038/nature05457.
- Schiffbauer, J. D., Xiao, S., Sharma, K. S. und Wang, G.: The origin of intracellular structures in Ediacaran metazoan embryos. In: Geology. Band 40 (3), 2012, S. 223–226, doi:10.1130/G32546.1.
- Chuanming, Z. u. a.: The diversification and extinction of Doushantuo-Pertatataka acritarchs in South China: causes and biostratigraphic significance. In: Geological Journal. Band 42, 2007, S. 229–262, doi:10.1002/gj.1062.
- S. Xiao und A. H. Knoll: Fossil preservation in the Neoproterozoic Doushantuo phosphorite Lagerstatte, South China. In: Lethaia. Band 32(3), 1999, S. 219–240.