River-Barrier-Hypothese
Die River Barrier Hypothesis (auch: Riverine Hypothesis, dt.: These, dass Flüsse Artbarrieren bilden; Flussbarrieren-Hypothese) ist eine wissenschaftliche Hypothese, die die hohe Biodiversität im Amazonasbecken und anderen Regenwaldgebieten erklären soll. Sie wurde zuerst von Alfred Russel Wallace in seiner Studie von 1852 On Monkeys of the Amazon (Über Affen des Amazonas) vorgestellt.[1] Die Theorie besagt, dass Lauf und Strömung des Amazonas und einiger seiner Nebenflüsse groß genug seien, dass sie als Hindernisse wirken und damit bei einigen Tierpopulationen allopatrische Artbildung herbeiführen. Aufgrund von abweichendem Selektionsdruck und Gendrift sowie fehlendem Genaustausch entwickeln sich die Populationen auf beiden Seiten auseinander und die Art spaltet sich auf.

Mehrere Indizien sollten zu beobachten sein, wenn Artbildung durch eine Flussbarriere verursacht wurde:
- Die Artunterschiede auf beiden Seiten des Flusses sollten mit zunehmender Breite des Flusses ebenfalls zunehmen und nur wenig oder gar nicht im Quellgebiet feststellbar sein.
- Organismen, die ausschließlich auf oder über festem Boden (Terra Firme) leben, sollten stärker betroffen sein als solche, die in den Auwäldern (Überschwemmungsgebieten) entlang des Flusses vorkommen, da die ersten eine längere Strecke überwinden müssen, bevor sie ein wieder passendes Habitat erreichen, und getrennte Tieflandpopulationen andererseits relativ häufig wieder zusammengeführt werden können, sobald ein Flusslauf sich in den frühen Stadien der Altwasserbildung (oxbow lake) verengt oder seinen Lauf verlegt.
- Wenn ein Flusslauf als Barriere die Teilung von Arten begünstigt, sollten Schwester-Arten auf den gegenüberliegenden Ufern häufiger auftreten, als man es bei einer zufälligen Verteilung erwarten würde.
Mechanismen
Eine Artbildung durch eine Flussbarriere erfolgt, wenn ein Fluss breit genug ist, um allopatrische Artbildung (Artbildung durch räumliche Trennung, mit vikarianter Verbreitung der Tochterarten) auszulösen, oder wenn der Fluss groß genug ist, um genetischen Austausch (Genfluss) zwischen zwei Populationen auf den gegenüberliegenden Ufern zu verhindern. Eine genetische und morphologische Auseinanderentwicklung der Population einer Art, deren Individuen ihn nicht überqueren können, wird der Hypothese gemäß eingeleitet, wenn der Fluss seinen Lauf sekundär in ihr Habitat hinein verlegt, oder wenn sich die Art auf beiden Seiten des Flusses von der Quelle zur Mündung hin ausbreitet, wodurch ihre Population am Unterlauf geteilt wird. Durch einen Zufall (etwa auf Treibgut schwimmend) könnte auch einmalig eine kleine Gründerpopulation einer anfangs nur auf einer Seite heimischen Art über den die gewöhnliche Ausbreitung verhindernden Fluss hinweg transportiert werden.
Gewöhnlich ist die Wahrscheinlichkeit, dass ein Fluss zur Barriere wird, proportional zu seiner Breite; je größer die Distanz, die zu überwinden ist, desto größer die Schwierigkeit, dass weiterhin Genaustausch stattfindet. Die Stärke der Barrierewirkung verändert sich im Verlauf eines Flusses; die schmalen Quellbäche sind leichter zu überqueren als die weiten Flussbetten im Flachland. Flüsse, die in einer Region für bestimmte Arten eine Barriere bilden, stellen für andere Arten kein Hindernis dar. Die Wanderungsbarriere ist also von Art zu Art (oder allgemeiner Klade zu Klade) unterschiedlich und trennt je nachdem unterschiedlich stark; Isolation und Differenzierung auf den gegenüberliegenden Ufern zeigen ein spezifisches Muster.
Große Säugetiere und Vögel haben nur wenig Schwierigkeiten, die großen Ströme zu überqueren, während kleine Vögel, die nur kurze Flugstrecken zurücklegen können, und kleine Säugetiere wahrscheinlich weniger häufig große Strecken überwinden und bei ihnen häufiger eine Auswirkung sichtbar sein sollte. Zudem ist bekannt, dass es zahlreiche Vogelarten vermeiden, über offene Wasserflächen zu fliegen. Flüsse können durch Mäanderbildung die zu überwindende Strecke für Tiere, die trockenere Wälder bevorzugen, vervielfachen. Umgekehrt kann die Abschnürung von Mäanderschlingen als Altwassern in Überschwemmungsgebieten Landgebiete im Inneren der Schlinge, mit ihren Besiedlern, von einem Ufer des Flusses zum anderen verfrachten.[2]
Stützende Indizien
Viele Forschungsprojekte im Amazonasbecken hatten das Ziel, die Gültigkeit dieser Hypothese zu überprüfen. Der Südliche Rotschwanz-Ameisenvogel (Sciaphylax hemimelaena) ist eine Art, die die Hypothese in der Natur bestätigt. Die Diversifikation und Verteilung des Ameisenvogels wurde im Amazonasgebiet untersucht. Dabei konnten drei monophyletische, genetisch abgrenzbare Populationen ausgemacht werden; zwei davon sind mittlerweile als Unterart anerkannt. Auf beiden Seiten des Rio Madeira kommen zwei der Kladen vor und die dritte hatte ihren Lebensraum zwischen dem Rio Madeira und zwei Zuflüssen, dem Jiparaná und dem Rio Aripuanã.[3] Die Untersuchungen weisen darauf hin, dass diese Vogelart sich durch Flussbarrieren aufgespalten hat, indem der Genfluss unterbrochen wurde.

Eine andere Studie an Sattelrückentamarinen (Leontocebus sp.) untersuchte die Voraussage, dass der Genfluss an verschiedenen Flussabschnitten unterschiedlich verläuft. Dabei wurde tatsächlich festgestellt, dass der Genfluss vor allem über die schmaleren Quellflüsse stattfindet, während er zur Mündung hin immer weiter abnimmt.[4] Auch dies stimmt mit der Hypothese überein. Jedoch ist fraglich, ob ein einziger Mechanismus ausreichend ist, die Diversifikation in den Tropen zu erklären. Beispielsweise gibt es Hinweise darauf, dass genetische Variation beim Blauscheitelpipra (Lepidothrix coronata) nur durch eine Kombination durch Flussbarrieren, Anden-Auffaltung und Ausbreitung der Art erklärbar sind.[5]
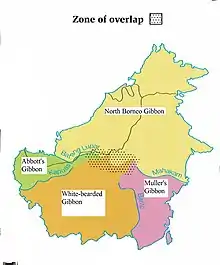
In der Primatologie wird die River-Barrier-Hypothese weitgehend akzeptiert. So teilte die Regenwaldexpertin und Primatologin Laura K. Marsh die Sakis (Pithecia) in einer Revision der Affengattung in 16 Arten ein und ging bei ihrer Arbeit davon aus, dass die großen Flüsse Amazoniens gemäß der River-Barrier-Hypothese nicht von den Affen überquert werden können und somit als Barrieren für eine allopatrische Artbildung wirken.[6] Ähnliche Verbreitungsmuster zeigen sich bei den Totenkopfaffen (Saimiri), z. B. im östlichen Amazonasbecken, wo der Gewöhnliche Totenkopfaffe nördlich des Amazonas vorkommt und der sehr ähnliche Collins-Totenkopfaffe südlich des Amazonas lebt,[7] bei den Ungehaubten Kapuzinern (Cebus), den Bartsakis (Chiropotes), den amazonischen Saguinus-Arten und bei den vier Gibbonarten (Hylobates sp.) auf Borneo.
Kritik

Nicht alle Studien erbrachten positive Beweise für die Hypothese. Eine Studie überprüfte die Riverine Hypothesis anhand der Populationen von vier Arten von Fröschen entlang des Rio Juará. Das Team erwartete Genfluss in geringerem Ausmaß, wenn es die Populationen auf gegenüberliegenden Ufern mit solchen verglich, die auf demselben Ufer lebten. Dies war jedoch nicht der Fall. Genfluss schien in beiden Fällen in gleich großem Ausmaß erfolgt zu sein.[8]
Eine andere Studie überprüfte die Hypothese in einem größeren Zusammenhang. Dabei gingen die Forscher von der These aus, dass Flüsse nicht nur Barrieren für den Genfluss innerhalb bestimmter Arten darstellen, sondern sogar auf Ebene ganzer Artengemeinschaften. Dazu wurde die Variation bei Froscharten und kleinen Säugetieren entlang von Flüssen und auf den gegenüberliegenden Ufern des Juará River untersucht. Dabei wurde keine sichtbare Abweichung zwischen den Froscharten und Säugetierarten an den Quellflüssen und an der Flussmündung festgestellt. Und es wurden auch keine größeren Ähnlichkeiten zwischen den Arten festgestellt, die auf demselben Ufer des Flusses leben, als zwischen Arten die auf gegenüberliegenden Uferseiten vorkommen.[9]
Diese Resultate stellen die Hypothese teilweise in Frage. Die Gültigkeit der Hypothese wurde daraufhin durch Studien an Baumsteigerfröschen weiter auf die Probe gestellt.[10] Dabei ergab sich, dass die Populationen auf derselben Seite des Flusses durchaus nicht alle monophyletisch verwandt waren. In der Studie von Lougheed sollte gezeigt werden, dass die Ridge Hypothesis (Gebirgskamm-Barrieren-Theorie) viel wahrscheinlicher ist, als die untersuchte Hypothese.[11]
81 Arten von flugunfähigen Säugetieren wurden in einem weiteren Experiment auf gegenüberliegenden Ufern des Rio Juará in Fallen gefangen. Der Fluss schien nur für wenige Arten eine Barriere zu sein, während der Großteil der Arten entweder im Untersuchungsgebiet homogen oder in monophyletische Kladen zwischen Oberlauf und Unterlauf unterteilt war. Patton argumentiert, dass die geographische Verteilung dieser Kladen eher darauf schließen lässt, dass Landform-Evolution (also Aufspaltung nach Geländehöhe, Untergrund etc.) ein unterschätzter Faktor der Diversifikation in Amazonien sei.[12]
Diese Untersuchungen sprechen dafür, dass auch andere Faktoren Einfluss auf die Artentstehung in Amazonien nehmen. Ein Mangel der Hypothese ist auch, dass sie fast ausschließlich in Amazonien untersucht wurde und noch kaum in anderen Strombecken. Außerdem ist es sehr wahrscheinlich, dass regelmäßige Verlagerungen des Flussverlaufs die Bildung von erkennbaren Mustern verhindern und es aus diesem Grund sehr schwierig ist, die Glaubwürdigkeit der Hypothese mit wissenschaftlichen Untersuchungen zu überprüfen.
Einzelnachweise
- A. R. Wallace: On the monkeys of the Amazon. In: Journal of Natural History, 1854, 14 (84): 451-454.
- vgl. dazu (am Mississippi in Nordamerika untersucht): Nathan D. Jackson, Christopher C. Austin (2013): Testing the Role of Meander Cutoff in Promoting Gene Flow across a Riverine Barrier in Ground Skinks (Scincella lateralis). PLoS ONE 8(5): e62812. doi:10.1371/journal.pone.0062812.
- Alexandre M. Fernandes, Michael Wink, Alexandre Aleixo: Phylogeography of the chestnut-tailed antbird (Myrmeciza hemimelaena) clarifies the role of rivers in Amazonian biogeography. In: Journal of Biogeography. 39 (2012): 1524-535.
- C. A. Peres, J. L. Patton, Maria Nazareth F. Da Silva: Riverine barriers and gene flow in Amazonian saddle-back tamarins. In: Folia Primatologica. 67.3 (1996): 113-24.
- Z. A. Cheviron, S. J. Hackett, A. P. Capparella: Complex evolutionary history of a Neotropical lowland forest bird (Lepidothrix coronata) and its implications for historical hypotheses of the origin of Neotropical avian diversity. In: Molecular Phylogenetics and Evolution. 36.2 (2005): 338-357.
- Laura K. Marsh (2014): A Taxonomic Revision of the Saki Monkeys, Pithecia Desmarest, 1804. Neotropical Primates. 21(1); 1-163.
- M. P. Mercês, Jessica W. Lynch Alfaro, W. A. S. Ferreira, M. L. Harada, José de Sousa e Silva Júnior: Morphology and mitochondrial phylogenetics reveal that the Amazon River separates two eastern squirrel monkey species: Saimiri sciureus and S. collinsi. Molecular Phylogenetics and Evolution, 82, Part B, S. 426–435, Januar 2015 (online ab 20. Oktober 2014) doi:10.1016/j.ympev.2014.09.020
- Claude Gascon, Stephen C. Lougheed, James P. Bogart: Patterns of genetic population differentiation in four species of Amazonian frogs: a test of the riverine barrier hypothesis. In: Biotropica 30.1 (1998): 104-19.
- Claude Gascon, Jay R. Malcolm, James L. Patton, James P. Bogart: Riverine barriers and the geographic distributions of Amazonian species. In: Proceedings of the National Academy of Sciences. 97.25 (2000): 13672-3677.
- Stephen C. Lougheed, Claude Gascon, D. A. Jones, James P. Bogart, Peter T. Boag: Ridges and rivers: a test of competing hypotheses of Amazonian diversification using a dart-poison frog (Epipedobates femoralis). In: Proceedings of the Royal Society. 266.1421 (1999): 1829-1835.
- Stephen C. Lougheed, Claude Gascon, D. A. Jones, James P. Bogart, Peter T. Boag: Ridges and rivers: a test of competing hypotheses of Amazonian diversification using a dart-poison frog (Epipedobates femoralis). In: Proceedings of the Royal Society. 266.1421 (1999): 1829-1835.
- James L. Patton, Maria Nazareth F. Da Silva, Jay R. Malcolm: Mammals of the Rio Juruá and the evolutionary and ecological diversification of Amazonia. In: Bulletin of the American Museum of Natural History 244 (2000): 1-306
Literatur
- G. Voelker, B. D. Marks, C. Kahindo, U. A'genonga, F. Bapeamoni, L. E. Duffie, J. W. Huntley, E. Mulotwa, S. A. Rosenbaum, J. E. Light: River barriers and cryptic biodiversity in an evolutionary museum. In: Ecology and evolution. Band 3, Nummer 3, März 2013, S. 536–545, doi:10.1002/ece3.482, PMID 23532272, PMC 3605844 (freier Volltext).
- R. K. Colwell: A barrier runs through it ... or maybe just a river. In: Proceedings of the National Academy of Sciences. 97, 2000, S. 13470, doi:10.1073/pnas.250497697.