Haarzelle
Haarzellen oder Haarsinneszellen sind ein Typ von sekundären Sinneszellen (Rezeptoren) im Nervensystem von Wirbeltieren, die mechanische Reize in Nervenaktivität umwandeln. Sie tragen apikal Stereozilien und gehören damit zur Klasse der Mechanorezeptoren. Haarzellen können je nach Typ durch Schall, Wasserströmungen, Dreh- oder Linearbeschleunigung erregt werden. Am besten untersucht sind die Haarzellen des Innenohres der Säugetiere.
Aufbau
Haarzellen bestehen aus dem Zellkörper und den namensgebenden haarähnlichen Strukturen, die zur Aufnahme des Reizes dienen. Dieses Haarbündel sitzt der Zelle am oberen Ende auf und besteht aus einer Zilie (Kinozilie) und mehreren Stereovilli. Beim Menschen werden die Zilien der Haarzellen der Hörschnecke im Gegensatz zu denen des Gleichgewichtsapparats nach der Geburt wieder zurückgebildet. Die einzelnen Stereovilli sind an den Spitzen miteinander verbunden, diese Verbindungen nennt man „Tip-Links“. Am unteren, dem Haarbündel entgegengesetzten Ende der Zelle befindet sich eine Region, in der die Erregung der Haarzelle zu einer Ausschüttung von Botenstoffen, den Neurotransmittern, führt. Hier bilden Haarzellen Synapsen mit Interneuronen, die die Information in Form von Aktionspotentialen weiter in das zentrale Nervensystem (ZNS) tragen.
Funktion
Reizaufnahme – Transduktion

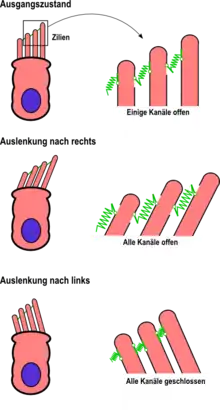
Die für die Reizaufnahme entscheidende Struktur der inneren Haarzellen bildet das Haarbündel. Die einzelnen Stereovilli sind an den Spitzen durch die Tip-Links verbunden. Am unteren Ende dieser Verbindungen (am kürzeren Stereovillus) befindet sich der Ionenkanal, der sogenannte Transduktionskanal, der je nach Spannung durch den Tip-Link geöffnet oder geschlossen wird. Allerdings konnte das Molekül, welches den Transduktionskanal bildet, noch nicht identifiziert werden.[1] Die Öffnung der Kanäle führt zu einem Einstrom positiver Kaliumionen, die die Zelle damit depolarisieren. Ohne eine auslenkende Kraft, die auf das Haarbündel wirkt, sind die Kanäle nur teilweise geöffnet – die Zelle ist also in Ruhe mittelmäßig erregt. Bei Auslenkungen der Stereovilli in Richtung der Zilien werden die Kanäle geöffnet und führen über den Einstrom des Kaliums zu einer Erregung der Haarzelle. Auslenkungen entgegen der Zilie schließen die Kanäle. Bewegungen auf einer anderen Achse als der durch Zilienanordnung bestimmten führen nicht zu einer Veränderung der Kanalöffnung und spielen damit keine Rolle für den Erregungszustand der Zelle.
Weiterleitung der Erregung
Im Gegensatz zu primären Sinneszellen kommt es bei den Haarsinneszellen (sekundäre Sinneszellen) nicht zur Bildung eines Aktionspotentials. Dieses könnte mit der hohen Geschwindigkeit des akustisch bewegten Rezeptors gar nicht generiert werden. Die Menge der ausgeschütteten Transmitter wird vielmehr von der Höhe des Rezeptorpotentials bestimmt.
Transduktionsmechanismus der Haarzellen im Innenohr
In der Cochlea des menschlichen Innenohres finden sich drei Reihen von äußeren und eine Reihe von inneren Haarzellen. Die sensorische Aufnahme mechanischer Bewegungen in der Cochlea erfolgt fast ausschließlich durch die inneren Haarzellen, während die äußeren Haarzellen v. a. efferente Innervierung durch übergeordnete Zentren des ZNS erhalten. Prinzipiell erfolgt die Transduktion der mechanischen Auslenkung der (inneren) Haarzellen im Innenohr in ein elektrisches Signal wie oben beschrieben durch Kaliumioneneinstrom. Es gibt jedoch einige Besonderheiten.
Ionenverteilung
Der untere, basale Teil der Haarzelle ist von Corti-Lymphe umgeben, die sich im inneren und äußeren Tunnel und dem Nuel-Raum des Corti-Organes befindet und die in ihrer Zusammensetzung der Perilymphe ähnlich ist – jener Flüssigkeit, welche die Scala vestibuli (und Scala tympani) füllt. Die Spitze der Haarzelle mit den Stereovilli befindet sich in der Endolymphe der Scala media. Die Perilymphe weist eine hohe Konzentration von Natrium- und eine niedrige Konzentration von Kaliumionen auf. In der Endolymphe ist dieses Verhältnis umgekehrt (viele Kaliumionen, wenig Natriumionen). Zwischen diesen beiden äußeren Bereichen der Haarzelle besteht ein Spannungsunterschied: die Endolymphe (oben) ist gegenüber der Perilymphe (unten) +85 mV positiv geladen. In Ruhestellung (wenn keine Auslenkung der Stereovilli erfolgt), ist das Zytoplasma der Haarzelle gegenüber der Perilymphe negativ geladen. Im oberen Teil der Haarzelle, der von der endolymphen Flüssigkeit umgeben ist, besteht zwischen dem Zelleninneren und der Umgebung ein Spannungsgefälle von −155 mV. Im unteren Zellbereich, welcher von der Perilymphe umgeben ist, besteht ein Spannungsunterschied zur Umgebung von −70 mV.[2]
Depolarisation
Werden die Stereovilli der Haarzellen durch mechanische Schwingungen der Basilarmembran der Cochlea in Richtung des längsten Stereoziliums ausgelenkt, bewirkt dies (wie oben beschrieben) über Tip-Link-Verbindungen die Öffnung von Kaliumkanälen in den Haarzellen. Im oberen Bereich der Haarzelle (Endolymphflüssigkeit) kommt es zu K-Ioneneinstrom. Dieser Einstrom kommt dadurch zustande, dass das Zelleninnere 155 mV negativer geladen ist als die Endolymphe. Dies führt dazu, dass positive Ladungen in Form von K-Ionen einströmen. Das chemische Gleichgewichtspotential von Kalium beträgt 0 mV, da intrazellulär die gleiche Konzentration herrscht wie in der Endolymphe[3], ist aber durch das elektrische Potential von −155 mV „bestrebt“, die Spannungsdifferenz zwischen Zelläußerem und -innerem zu positivieren. Die Kaliumionen bewirken im Zellinneren die Öffnung von Calciumkanälen, wodurch Calcium einströmt. Dies führt wie in anderen Neuronen zur Depolarisation und damit zur verstärkten Ausschüttung von Neurotransmittern an nachgeschaltete Neurone.
Repolarisation
Die Besonderheit der Transduktion besteht darin, dass Kalium sowohl für die De- als auch für die Repolarisation zuständig ist. Die in den oberen Teil der Haarzelle eingeströmten Kaliumionen führen ihrerseits zur Öffnung weiterer Kaliumkanäle in der gesamten Zellmembran. Das durch die Depolarisation vermehrt vorhandene Calcium führt u. a. ebenfalls zur Öffnung von K-Kanälen. Im unteren, von Perilymphe umgebenen Zellbereich besteht jedoch mit −45 mV ein geringerer Spannungsunterschied zur Umgebung als im oberen Bereich. Das oben eingeströmte Kalium strömt über Kaliumkanäle im unteren Teil der Zelle wieder aus, da
- in der Perilymphe im Vergleich zum Zellinneren eine sehr geringe Kaliumkonzentration herrscht
- Kalium bestrebt ist, sein Gleichgewichtspotential von −80 mV herzustellen
Letzteres führt dazu, dass positive Ladungen in Form von K-Ionen ausströmen müssen, um die Spannungsdifferenz von −45 mV auf −80 mV zu senken. Durch den Kaliumausstrom kommt es zur Repolarisation der Haarzelle.
Motorfunktion der Haarbündel

Neuere Forschungsergebnisse zeigten, dass die Bündel der Stereozilien (Haarbündel) im Innenohr der Landwirbeltiere neben ihrer Funktion als Mechanorezeptoren auch die Funktion von Motoren (Motilität) haben. Hierbei arbeiten die mechano-elektrischen Wandler in den Enden der Stereozilien in umgekehrter Richtung, also als elektro-mechanische Wandler. Sie geben dabei Energie ab und verstärken damit die Schallwellen, von denen sie angeregt werden. Diese Identität von Sensor- und Motorfunktion der Stereozilien dient nach bisherigen Hypothesen der Verbesserung der Frequenz-Abstimmung und damit der Frequenzauflösung des Hörorgans. Bei Nicht-Säugern ist diese Hypothese inzwischen bewiesen und allgemein anerkannt. Bei Säugern, wo es zusätzlich die Sondererscheinung der Motorfunktion auch der Zellkörper der äußeren Haarzellen gibt (Cochleärer Verstärker), ist noch nicht geklärt, wie im Einzelnen Haarbündelmotoren und Zellkörpermotoren zusammenwirken.[4]
Quellen
- David P. Corey (2006): What is the hair cell transduction channel?, J. Physiol. 576: 23–28.
- Zenner H.-P.: Hören. Physiologie, Biochemie, Zell- und Neurobiologie. G. Thieme Verlag, Stuttgart, 1994.
- Klinke, Silbernagl, Auflage 2005, S. 664
- James O. Pickles: An Introduction to the Physiology of Hearing, Bingley, Emerald Group Publishing 2012, 430 S. ISBN 1-78052-166-9, S. 135 und 137.
Weblinks
- Confocal Image of Cochlea Wins Art Prize. Retrospectacle: A Neuroscience Blog, 14. November 2007, abgerufen am 4. Januar 2014 (englisch, konfokale Abbildung der Cochlea mit Haarzellen).
- Journey into the World of Hearing. Association NeurOreille, abgerufen am 4. Januar 2014 (englisch).