Lepidodendron
Lepidodendron ist eine Gattung der „Schuppenbäume“, der baumförmigen Bärlappgewächse, die die Steinkohlenwälder des Erdzeitalter des Karbon wesentlich mit aufgebaut haben. Der Name ist eingeführt worden für ein Formtaxon, für Stammabschnitte, deren Abdrücke oder Steinkerne mit einer charakteristischen Struktur der Rinde. Er wurde später auf die auf deren Basis rekonstruierten Pflanzen übertragen, was aber nicht ganz unumstritten ist. Der Name Lepidodendron geht zurück auf das Werk Versuch einer geognostisch-botanischen Darstellung der Flora der Vorwelt des böhmischen Grafen und Begründers der Paläobotanik Kaspar Maria von Sternberg von 1820, dass nach dem nomenklatorischen Code der Botanik Ausgangspunkt für die Nomenklatur für fossile Organismen ist (ICN, Artikel 13). Damit ist es einer der ältesten Namen für einen fossilen Organismus, der auch heute noch in Gebrauch ist. Bäume der Gattung Lepidodendron erreichten 40 Meter Höhe bei einem basalen Stammdurchmesser von zwei Meter.
| Lepidodendron | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Stammabdruck von Lepidodendron mit den charakteristischen Abdrücken der Blattpolster | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Viséum bis Changhsingium | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Lepidodendron | ||||||||||||
| Sternb. |
Merkmale
Stammabdrücke
Die „Schuppenbäume“ des Karbon erhielten ihren Namen nach Bruchstücken von Stammabdrücken, die ein charakteristisches, regelmäßiges Schuppenmuster aufweisen. Später wurde erkannt, dass diese „Schuppen“ Blattpolster, d. h. die verbreiterte Blattbasis abgeworfener Laubblätter sind (in der Form der für die Bärlappgewächse charakteristischen Mikrophylle, die entgegen der Benennung aber durchaus beinahe einen Meter Länge erreichen konnten). Die Blattpolster sind bei Lepidodendron[1][2] ungefähr rhombisch, mit spitzen oberen und unteren Ecken und abgerundeten Seiten, dabei immer höher als breit. Die eigentliche Blattnarbe der abgeworfenen Blattspreite sitzt ein wenig oberhalb der Mitte der Struktur, sie ist elliptisch oder rhombisch. Auf ihrer Oberfläche sind bei besser erhaltenen Exemplaren drei kleine Marken erkennbar, deren mittlere auf die einzige Blattader des Mikrophylls zurückgeht. Die beiden seitlichen waren „Parichnos“ genannte Brücken parenchymatischen Gewebes, möglicherweise ein Luftleitgewebe (Aerenchym), die bei rezenten Pflanzen unbekannt und charakteristisch für die Lepidodendrales sind. Typisch für Lepidodendron sind zwei zusätzliche Parichnos auf den Abdrücken der Blattpolster wenig unterhalb der Blattnarbe; diese fehlen den Schuppenbäumen der Familie der Diaphorodendraceae. Von der Blattnarbe geht eine vertikale, kielartige Linie von der Blattnarbe zur oberen bzw. unteren Spitze des Blattpolsters, die so vier Felder, zwei größere unten und zwei kleinere oben, abgrenzt. Bei Lepidodenddron ist dieser Kiel zumindest im unteren Abschnitt etwas quer gefältelt. Außerdem laufen bei Lepidodendron, nicht aber bei den meisten anderen Schuppenbäumen, noch zwei sogenannte laterale Linien von der Blattnarbe horizontal zum Rand des Blattpolsters. Die vermutlich die Schwestergruppe von Lepidodendron bildende Gattung Lepidophloios ist von dieser leicht an den Blattpolstern unterscheidbar, die bei dieser merklich breiter als lang sind.
Rekonstruktion der ganzen Pflanzen
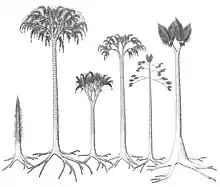
Die Bärlappbäume von Lepidodendron sind, wie typisch für fossile Bäume, in der Regel nur in einzelnen Organen oder deren Bruchstücken überliefert. Abgeworfene Blätter, Zweige und reproduktive Organe (Zapfen mit Sporophyllen), oder auch die Stammbasis mit den Wurzeln, sind meist getrennt von ihrer Mutterpflanze fossiliert und haben dem entsprechend zunächst eigene Namen, als Formtaxa, erhalten. Im Fall von umfassend und in vielen Tausend Exemplaren erhalteten Fossilien wie Lepidodendron war es, oft durch glückliche Zufallsfunde mit ausnahmsweise vollständiger erhaltenen Exemplaren, möglich, nach und nach die gesamte Pflanze aus den erhaltenen Bruchstücken zu rekonstruieren. Dies ist, in mehr oder weniger vollständiger und überzeugender Form, bei etwa 16 Arten von Bärlappbäumen gelungen. Die übrigen Formtaxa sind weiterhin nicht solchen rekonstruierten Arten zuzuweisen, so dass die tatsächliche Artenzahl unsicher bleibt. Es ist auch durchaus wahrscheinlich, dass die Belaubung und insbesondere das Wurzelwerk recht unspezifisch war, so dass auch im Falle erfolgreicher Rekonstruktionen immer die Möglichkeit besteht, dass ein Formtaxon mehr als einer Art entspricht.
Im Falle von Lepidodendron wurden zusätzlich zu den häufigen Stammabdrücken oder inkohlten Kompressionsfossilien Bruchstücke von permineralisiertem Holz als Lepidodendron hickii beschrieben, anhand derer die innere Morphologie der Stammachse rekonstruiert werden konnte.[3] Das bei den Bärlappbäumen Stigmaria genannte Wurzelwerk der Formart Stigmaria ficoides gehört ebenfalls dazu. Reproduktive Organe sind die Megasporen-bildenden Zapfen Achlamydocarpon takhtajanii, die Mikrosporen-bildenden Zapfen Lepidostrobus cf. oldhamius mit den dazugehörigen Sporen Lycospora pusilla, später dem Artaggregat Lycospora brevijuga zugeordnet.[2] Die, nicht unbedingt artspezifischen, isolierten Laubblätter sind als Lepidophylloides beschrieben.
Nach den fossilen Funden war Lepidodendron ein monokarper Baum, der nur einmal am Ende seines Lebenszyklus Sporen bildete und anschließend abstarb. Der Stamm bildete sich aus dem ausgedehnten, unter- bis oberirdisch kriechenden Stigmaria-„Wurzel“werks (Rhizomorph genannt), das auch photosynthetisch aktiv und assimilierend war. Der Stamm war zunächst völlig gerade säulenartig und unverzweigt (als Wuchsform „pole tree“, d. h. „Pfostenbaum“ genannt) und in den oberen Abschnitten mit direkt aus dem Stamm aus den Blattpolstern entspringenden Blättern bedeckt, eine entsprechende Wuchsform ist bei rezenten Bäumen nicht mehr bekannt. Vor der Bildung der sporentragenden Strobili verzweigte der Baum in einer lockeren, gabelteiligen Krone. Bei wenigen Arten waren zusätzlich, ebenfalls Sporophylle tragende, Seitenäste ausgebildet, die ebenfalls an einer präfomierten Zone abgeworfen wurden und dann charakteristische Astnarben hinterließen („ulodendroid“ genannt[4]). Die nach dem Abwurf der Blätter zurückbleibenden Blattpolster trugen zahlreiche Stomata und waren mit hoher Wahrscheinlichkeit photosynthetisch aktiv.[4] Der Stamm wäre also im Leben grün gefärbt gewesen.
Taxonomie und Systematik
Sternberg unterschied, nach Details der Blattbasen, sieben Arten von Lepidodendron, von denen Lepidodendron dichotomum, durch nachträgliche Festsetzung durch Samuel Almond Miller 1889, zur Typusart der Gattung erklärt wurde, was aber nicht immer beachtet wurde.[5] Das Typusexemplar (ein Lectotyp, gefunden in Tschechien) liegt im Nationalmuseum Prag. Von den verbleibenden Arten wurde eine in die Gattung Lepidophloios transferiert, eine ist wegen schlechter Erhaltung nicht zuzuordnen, die anderen vier wurden als konspezifisch erkannt und drei Artnamen unter Lepidodendrun aculeatum synonymisiert; diese Art betrachten andere Paläobotaniker als die valide Typusart.[3]
Arten
Es werden aktuell gut 30 Arten der Formgattung Lepidodendron benannt, die nach Details der Gestalt der Blattpolster, etwa der Position und Größe der Parichnos, unterschieden werden.[5] Vor den moderneren Bearbeitungen mit der Aufspaltung in zahlreiche Gattungen wurden zeitweise mehr als 400 Namen unter Lepiodendron geführt. Die meisten davon wurden später in andere Gattungen transferiert oder sind aufgrund von Merkmalsarmut oder schlechter Erhaltung nicht sicher zuzuordnen.[6]
Verbreitung
Lepidodendron gehörte zu den charakteristischen Elementen der ausgedehnten Torfwälder der tropischen Breiten im Karbon, aus deren Torf durch Inkohlung die Steinkohlenlagerstätten hervorgegangen sind. So bildeten Lepidodenron-Stämme bis zu drei Viertel der inkohlten Biomasse der Küstensümpfe des Mississippium in Nord-China.[7] Die Gattung im engeren Sinne war aber wohl auf Mineralböden häufiger als auf torfigem Untergrund, wo sie von anderen Bärlappbäumen wie vor allem Lepidophloios abgelöst wurde. Eine der Ausnahmen mit Verbreitungsschwerpunkt in den Torfwäldern ist die lange und gut bekannte Lepidodendron aculeatum (vermutlich identisch mit dem permineralisierten Fossil Lepidodendron hickii).[8]
Anhand der Untersuchung der Lagerstätten wird für Lepidodendron ein Vorkommen in extrem nassen, möglicherweise längere Zeit überschwemmten Habitaten rekonstruiert. Die flach wurzenden Bäume waren nicht imstande, Austrocknungsphasen zu überleben. Vermutlich waren sie auch für die Keimung der Sporen auf extrem nasse Böden angewiesen.
Der Formgattung Lepidodendron zugeordnete Fossilien sind fast weltweit in Steinkohlenlagerstätten des Karbon gefunden worden. Die ältesten fossilen Funde stammen aus dem frühen Karbon (Viséum).[8] Die zugehörigen Bäume sind, wie alle Bärlappbäume mit Ausnahme der Gattung Sigillaria, noch im Karbon in Europa und Nordamerika (damals ein einziger, Euramerika genannter Kontinent) ausgestorben. Sie überlebten auf den großen Inseln von Cathaysia (den, damals getrennte Inseln bildenden Nordchinakraton und Yangtze- oder Südchina-Kraton) bis ins mittlere Perm. In Nordamerika starb Lepidodendron etwa an der Grenze vom mittleren zum oberen Pennsylvanium (entspricht der Grenze vom Moskovium zum Kasimovium nach moderner Nomenklatur) aus.[8] Dasselbe ist für Europa zu beobachten[9], wo in den meisten Regionen im Stephanium gar keine Kohle mehr gebildet wurde. In China überlebte die Gattung, nach Funden aus der Kayitou Formation und der Xuanwei Formation, lokal bis unmittelbar zum Ende des Perm.[10]
Einzelnachweise
- Thomas N. Taylor, Edith L. Taylor, Michael Krings: Paleobotany. The Biology and Evolution of Fossil Plants. Second Edition, Academic Press 2009, ISBN 978-0-12-373972-8. S. 282–283.
- Richard M. Bateman & William A. DiMichele (2021): Escaping the voluntary constraints of “tyre-track” taxonomy. Taxon 70 (5): 1062–1077. doi:10.1002/tax.12540
- William A. DiMichele (1983): Lepidodendron hickii and Generic Delimitation in Carboniferous Lepidodendrid Lycopods. Systematic Botany 8(3): 317–333.
- Barry A. Thomas (1978): Carboniferous Lepidodendraceae and Lepidocarpaceae. Botanical Review 44 (3): 321-364.
- Barry A. Thomas & Christopher J. Cleal (2020): The nomenclature of fossil-taxa representing different preservational states: Lepidodendron as a case-study. Taxon 69 (5): 1052–1061. doi:10.1002/tax.12291
- Carmen Álvarez-Vázquez and Robert H. Wagner (2014): Lycopsida from the lower Westphalian (Middle Pennsylvanian) of the Maritime Provinces, Canada. Journal of the Atlantic Geoscience Society 50: 167-232. doi:10.4138/atlgeol.2014.011
- Jun Wang: Late Mississippian Coastal Vegetation in China. Chapter 13.3 in Edoardo Martinetto, Emanuel Tschopp, Robert A. Gastaldo: Nature through Time, Virtual field trips through the Nature of the past. Springer Verlag 2020. doi:10.1007/978-3-030-35058-1_13
- Tom L. Phillips, William A. DiMichele (1992): Comparative Ecology and Life-History Biology of Arborescent Lycopsids in Late Carboniferous Swamps of Euramerica. Annals of the Missouri Botanical Garden 79 (3): 560-588.
- William A. DiMichele and Tom L. Phillips (1994): Paleobotanical and paleoecological constraints on models of peat formation in the Late Carboniferous of Euramerica. Palaeogeography, Palaeoclimatology, Palaeoecology 106: 39-90.
- Zhuo Feng, Hai-Bo Wei, Yun Guo, Xiao-Yuan He, Qun Sui, Yu Zhou, Hang-Yu Liu, Xu-Dong Gou, Yong Lv (2020): From rainforest to herbland: New insights into land plant responses to the end-Permian mass extinction. Earth-Science Reviews 204: article 103153. doi:10.1016/j.earscirev.2020.103153