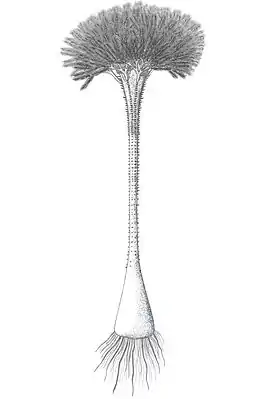
Calamophyton
Calamophyton ist eine ausgestorbene, wenige Dezimeter hohe Pflanze mit dichotomer Verzweigung. Sie gehört zur Gruppe der nahe der Basis der Farne stehenden Pseudosporochnales.
| Calamophyton | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
Calamophyton | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mitteldevon | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Calamophyton | ||||||||||||
| Kräusel & Weyland |
Merkmale
Calamophyton war mindestens 60 cm hoch.
Calamophyton primaevum, ursprünglich beschrieben aus dem Mitteldevon von Deutschland, wurde als 35 cm hohe Pflanze rekonstruiert mit fingerartig verzweigter Hauptachse, an der in schraubiger Anordnung segmentierte Blätter stehen. Die kurzen, dichotom verzweigten Zweige tragen an jedem Ende ein einzelnes Sporangium. Im Querschnitt zeigt sie eine variable Anzahl von Xylemarmen, die von 10 bis 33 reicht. Im oberen Teil der Pflanze gab es weniger Xylemarme. Die Xylemstränge sind im Querschnitt radial verlängert. Im Grundgewebe des Stammes befinden sich Faserbündel.
Calamophyton bicephalum aus dem Mitteldevon besitzt eine zentrale Achse mit fingerförmig angeordneten Seitenzweigen. Die Seitenzweige haben einen Durchmesser von rund 1 cm und verzweigen sich dichotom. Im Querschnitt besitzt die Achse 14 bis 16 Gefäß-Segmente, die entweder oval sind oder 4 bis 5 radiale Arme aufweisen. Es gibt nur primäres Xylem.
Die sterilen, endständigen Anhängsel werden von unten nach oben kleiner und weniger gut entwickelt. Die Blätter sind dreidimensionale Strukturen, jedes teilt sich dichotom einmal horizontal und einmal vertikal. Junge Blätter im distalen Bereich der Pflanze gabeln sich nur einmal, während sich die im unteren Bereich bis zu viermal gabeln.
Es gibt Hinweise, dass sich Blätter und Sporangien auf unterschiedlichen Zweigen befunden haben. Die Sporangien sind nicht zu Zapfen vereint. Die sporangientragenden Zweige stehen gegenständig entlang der fertilen Achse in pseudowirteliger Anordnung. Manchmal stehen sporangientragende Zweige auch zwischen sterilen Wirteln. Jeder fertile Ast besteht aus einem basalen Stiel, der in ein oberes und unteres Segment geteilt ist. Jedes Segment teilt sich weiter in drei kurze Seitenzweige, die jeweils in einer kurzen Gabel enden, an deren Ende je ein einzelnes Sporangium steht. Derart trägt bei Calamophyton bicephalum jeder Zweig 12 Sporangien. Die Sporangien sind zylindrisch und am Ende zugespitzt. Sie dürften sich an der ventralen Seite (Unterseite) geöffnet haben. Die Sporen von Calamophyton bicephalum reichen in der Größe von 86 bis 166 µm. Jede Spore ist im Umriss kreisförmig und hat eine deutliche trilete (dreistrahlige) Narbe, an der Oberfläche stehen zierliche Stacheln.
Die zunächst als Hyenia beschriebenen Funde werden heute ebenfalls zur Gattung Calamophyton gerechnet. Die ursprünglich als Rhizome interpretierten Funde werden heute als Serien von Seitenzweigen interpretiert. Von den Zweigen gehen dichotom verzweigte Anhängsel ab, von denen jedes drei Paare von länglichen Sporangien besitzt. Die Sporangien öffnen sich mit Längsschlitz, die Sporen sind groß und trilet. Die sterilen Anhängsel sind dreidimensionale Gebilde, die sich mehrmals dichotom gabeln.
Ein Exemplar von Hyenia elegans, das Schweitzer 1972 beschrieb, besteht aus einem fast 2 m langen Rhizom von 4,4 cm Durchmesser. An ihm stehen in schraubiger Anordnung Seitenäste, die sich dichotom verzweigen.
Systematik und Botanische Geschichte
Heute wird Calamophyton aufgrund der Achsenanatomie zu den Pseudosporochnales gestellt. Ursprünglich wurden mehrere Gattungen bzw. Arten unterschieden, die nach der Arbeit von Farin-Demaret und Berry (2000) jedoch alle zu Calamophyton primaevum gehören. Manche Autoren differenzieren weiterhin zwischen Calamophyton primaevum und Calamophyton bicephalum.
Calamophyton und Hyenia, beide aus dem Devon, wurden 1926 von Kräusel und Weyland in die Gruppe der Protoarticulatae (= Hyeniales) gestellt, eine heute obsolete Gruppe devonischer krautiger Pflanzen, die nach ihren Merkmalen Ähnlichkeiten mit den Sphenophyta aufweisen. Sie wurden als dichotom verzweigt und aus einem Rhizom heraus entstehend interpretiert. Die letzten Zweige hatten sterile Anhängsel, die in Pseudowirteln stehen. Sporangien stehen an abgewandelten Zweigen. Die Hyeniales sollten eine evolutionäre Parallelentwicklung zu den devonischen Lycophyten, etwa den Drepanophycales und Protolepidodendrales darstellen.
Durch Funde von versteinerten Achsen von Calamophyton (Leclercq und Schweitzer 1965, Schweitzer 1972, 1973) wurde offenbar, dass diese Gattung eine einmalige Anatomie hat, die in keiner Weise den Sphenophyta ähnelt.
Hyenia elegans wie auch Cladoxylon scoparium werden heute in das Artkonzept von Calamophyton primaevum einbezogen. Da alle drei Arten von Kräusel und Weyland 1926 in derselben Publikation erstbeschrieben wurden, ist die Frage des gültigen Namens komplex. Fairon-Demaret und Berry schlugen dafür Calamophyton primaevum vor, aus folgenden Gründen: das Typus-Material von Hyenia aus Norwegen konnte nicht beurteilt werden und die Anatomie passt nicht in das Gattungskonzept von Cladoxylon, zumal alle anderen Arten von Cladoxylon aus dem Mississippium stammen.
Belege
- Thomas N. Taylor, Edith L. Taylor, Michael Krings: Paleobotany. The Biology and Evolution of Fossil Plants. Second Edition, Academic Press 2009, ISBN 978-0-12-373972-8, S. 396–398.