Retinale Ganglienzelle
Als Ganglienzellen der Netzhaut oder retinale Ganglienzellen (RGC[1]) werden verschiedene in der Ganglienzellschicht Stratum ganglionare der Retina gelegene Nervenzellen des Auges bezeichnet, deren Axone zusammen den Sehnerv bilden.
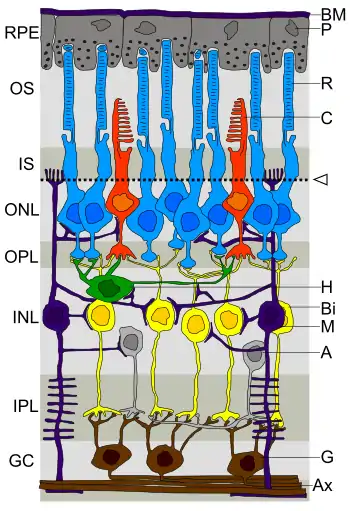
R: Stäbchen, C: Zapfen,
H: Horizontalzelle, Bi: Bipolarzelle,
A: Amakrinzelle, G: Ganglienzelle,
GC: Ganglienzellschicht.
(Das Licht fällt hierbei von unten ein.)
Im visuellen System stellen retinale Ganglienzellen das dritte afferente Neuron der Sehbahn dar: Von zwischengeschalteten Neuronen wie den Bipolarzellen als zweitem afferentem Neuron empfangen sie aufgearbeitete Informationen über die Erregung der Lichtsinneszellen – Stäbchen oder Zapfen als erstes afferentes Neuron –, verarbeiten diese Information und leiten daraufhin unterschiedliche eigene Signale an Kerngebiete des Corpus geniculatum laterale (CGL) im Metathalamus des Zwischenhirns weiter. Von dort zieht die Sehstrahlung als Projektionsbahn zum primären visuellen Cortex.
Im Sehnerv laufen weniger zahlreich auch Axone von retinalen Ganglienzellen, deren Signale für grundlegende Funktionen wie den Tag-Nacht-Rhythmus oder für optische Reflexe gebraucht werden. Diese Axone enden an
- Kernen des Nucleus suprachiasmaticus im Hypothalamus (Licht als Zeitgeber circadianer Rhythmen),
- Kernen der Area pretectalis im Epithalamus (afferenter Reflexbogenanteil der Pupillenlichtreaktion),
- Kernen der oberen Hügel im Mittelhirn (Afferenzen für reflektorische Augenbewegungen).
Morphologie
Die Zellkörper der Ganglienzellen liegen in der innersten, dem Glaskörper zugewandten Zellschicht (Stratum ganglionare) der Netzhaut. Nur im optischen Teil der Retina findet man diese Ganglienzellschicht (GC). Am stärksten ausgeprägt ist sie im gelben Fleck (Macula lutea) um die Netzhautmitte herum, wird zur Mitte hin dünner und fehlt in der zentralen Sehgrube (Fovea centralis). Denn an dieser Stelle sind die andernorts bedeckenden Innenschichten seitwärts verlagert und so die Sinneszellen der äußeren Körnerschicht (ONL) dem einfallenden Licht freigelegt; hier sind ausschließlich Zapfen zu finden, und in der Foveola nur M- und L-Zapfen. Die für diese zentralen Netzhautregionen zuständigen Ganglienzellen befinden sich in den umgebenden Randzonen der Makula.
Die Dendriten retinaler Ganglienzellen verzweigen sich je nach Zelltyp unterschiedlich reich und bilden in der inneren plexifomen Schicht (IPL) Synapsen mit den Bipolarzellen und den Amakrinzellen der weiter außen gelegenen inneren Körnerschicht (INL).
Die Axone aller Ganglienzellen, bei einer menschlichen Retina bis anderthalb Millionen, laufen inwendig als Nervenfaserschicht (Stratum neurofibrarum) zum Sehnerv (Nervus opticus) zusammen und verlassen an der Papille (Discus nervi optici) den Augapfel. Im Gesichtsfeld entspricht dieser Stelle der blinde Fleck. Von dort ziehen die Axone über das Chiasma opticum, teilweise kreuzend, überwiegend zu Kerngebieten der seitlichen Kniehöcker (Corpora geniculata lateralia, CGL) im Zwischenhirn, wo sie Synapsen mit nachgeschalteten Neuronen bilden.
Retinale Ganglienzellen sind multipolare Neurone mit langem Axon, die in Größe, Bau und Verzweigungsmuster verschieden sein können. Allein nach morphologischen Kriterien – beispielsweise nach der Erscheinungsform ihres Dendritenbaums als „winzig (midget)“, „spärlich (sparse)“, „dornig (thorny)“ oder „zweischichtig (bistratified)“ geschieden – gibt es über ein Dutzend Typen.[2]
Funktionelle Zuordnung
Darüber hinaus werden Ganglienzellen nach den Zielorten der Projektion aufgefächert, anhand elektrophysiologischer Kenndaten wie der Leitungsgeschwindigkeit klassifiziert, sowie hinsichtlich des Spektrums von Lichtreizen und der ausgelösten Signalmuster gekennzeichnet. Damit erst lassen sich dann mehrere Typen so differenzieren, dass ihnen unterschiedliche Aufgaben bei der Informationsverarbeitung zugeschrieben werden können.
Die Ganglienzellen sind die einzigen Zellen der Netzhaut, die Aktionspotenziale ausbilden können; bei den übrigen retinalen Neuronen ist die Erregungsleitung elektrotonisch. Fortgeleitet werden diese Aktionspotenziale im Auge über die als innerste Netzhautschicht verlaufenden Nervenfasern. Bei Primaten wie dem Menschen sind diese marklos ausgebildet, was nur eine relativ langsame Weiterleitung erlaubt – doch die Lichtbrechung verringert. Mit Austritt aus dem Auge werden die einzelnen Axone der Ganglienzellen dann als Sehnervenfasern markreich von einer Myelinscheide umgeben, wodurch eine schnellere Leitung der Signale möglich wird. Der Sehnerv insgesamt ist von Fortsetzungen der Hirnhäute umhüllt und so auch anatomisch als Teil des Hirns zu erkennen.
Rezeptive Felder
Der Bereich der Netzhaut, der Einfluss auf den Erregungszustand einer bestimmten Ganglienzelle nehmen kann, wird als das ihr zugeordnete rezeptive Feld bezeichnet. Ein rezeptives Feld umfasst eine bestimmte Gruppe von Rezeptorzellen – Stäbchen oder Zapfen – und entsteht durch die Konvergenz mehrerer Bipolarzellen auf eine Ganglienzelle. Horizontalzellen und Amakrinzellen wirken bei der Informationsweitergabe mit, vor allem durch laterale Hemmung.
Meist lassen sich rezeptive Felder in ein Zentrum und dessen Peripherie unterteilen. Diese beiden Bereiche haben eine gegensinnige Wirkung auf die Ganglienzelle. Ist das Zentrum erregend und die Peripherie hemmend, so spricht man von einer ON-Zentrum-Ganglienzelle, im umgekehrten Fall von einer OFF-Zentrum-Ganglienzelle. Mit solch einer Form der Verschaltung kann unter anderem der Kontrast einer visuellen Wahrnehmung verstärkt werden. Beispielsweise ist die Aktionspotenzial-Frequenz einer ON-Zentrum-Ganglienzelle dann besonders hoch, wenn die zugeordneten Fotorezeptoren des Zentrums sehr stark und die in der Peripherie ziemlich wenig gereizt werden.[3]
Die Größe der einzelnen rezeptiven Felder ist recht unterschiedlich. Sie hängt sowohl von dem Typ der Ganglienzelle als auch von ihrer Positionierung in der Netzhaut ab. Innerhalb der Macula lutea sind die rezeptiven Felder typischerweise ausgesprochen klein und umfassen nur wenige Zapfen. Für die Fovea centralis als den Bereich des schärfsten Sehens – und hier insbesondere für die Foveola als Bezugsstelle "Mitte" der räumlichen Relationen auf der Netzhaut – findet sich schließlich auch eine Konvergenz von 1:1. Außerhalb der Makula umfassen die rezeptiven Felder deutlich mehr Sinneszellen und nehmen zur Netzhautperipherie hin an Größe zu.
Zelltypen und Informationsverarbeitung
Nach dem gegenwärtigen Stand des Wissens werden bei der visuellen Informationsverarbeitung in Primaten drei Haupttypen retinaler Ganglienzellen unterschieden:[4]
- parasol-Ganglienzellen – Auf diesen Zelltyp konvergieren so genannte diffuse Bipolarzellen, die ihrerseits ihren Input von L- und M-Zapfen erhalten, also jenen Sinneszellen, die besonders empfindlich auf lang- bzw. mittelwelliges Licht reagieren. Die rezeptiven Felder sind vergleichsweise groß und die Dendritenbäume stark ausgebildet (parasol, englisch für ‚Sonnenschirm‘). Da die parasol-Ganglienzellen offensichtlich nicht zwischen den beiden Zapfentypen unterscheiden, sind sie "unfarbig", ihr Ausgangssignal ist achromatisch. Es dient vermutlich hauptsächlich der Hell-Dunkel-Unterscheidung und wird an die magnozellulären Schichten des CGL weitergegeben. Parasol-Ganglienzellen verarbeiten daneben auch Signale von Stäbchen, die mit ihrer hohen Sensitivität vor allem dem Dämmerungssehen dienen und zwischen verschiedenen Spektralbereichen nicht unterscheiden.
- midget-Ganglienzellen – Im Vergleich zu den parasol-Zellen haben die midget-Ganglienzellen einen kleinen Dendritenbaum und winzige rezeptive Felder, deren Zentrum meist einzig nur einen M-Zapfen oder einen L-Zapfen umfasst (midget, englisch für ‚Winzling‘). Dieses über midget-Bipolarzellen zugeordnete Zentrum kann ON oder OFF geschaltet sein, die umgebende Periphere dann jeweils umgekehrt. Im gemeinsamen Zusammenspiel leisten sie die Aufarbeitung des Rot/Grün-Kontrasts und verrechnen die Signale von L-Zapfen und M-Zapfen gegeneinander als Differenz (vereinfacht: L − M oder M − L). Die Axone der midget-Ganglienzellen projizieren in die parvozellulären Schichten des CGL. Bei diesem Teilsystem handelt es sich um das evolutionär jüngste; die unterschiedlichen Opsine der M- und L-Zapfen sind erst bei den Primaten durch eine Genduplikation entstanden[5]. In der menschlichen Retina sind etwa 80 % der Ganglienzellen vom midget-Zelltyp.
- bistratified-Ganglienzellen – Dieser Zelltyp hat sehr große rezeptive Felder mit einem ON-Zentrum von mehreren S-Zapfen, also den für kurzwelliges Licht besonders empfindlichen Sinneszellen. Deren Signale werden über konvergierende blue-Bipolarzellen erregend an Dendriten der Ganglienzelle geleitet ("Blue-ON"). Eine Peripherie im eigentlichen Sinn fehlt hier; diffuse Bipolarzellen sammeln hingegen Signale von M- und L-Zapfen und geben sie hemmend (OFF) an eine zweite Dendritenschicht der bistratified-Ganglienzellen weiter (bistratified, für ‚zweifach geschichtet‘). Damit wird dem Signal der S-Zapfen ein kombiniertes Signal von L- und M-Zapfen gegenübergestellt und verrechnet (vereinfacht: S − (M+L)). Auf diese Weise kann der Blau/Gelb-Kontrast hervorgehoben werden. Die Axone der bistratified-Ganglienzellen ziehen in die koniozellulären Schichten des CGL, die als Unterschichten interlaminar jeweils einer groß- bzw. kleinzelligen Schicht anliegen.
Nach den Zelltypen ihrer jeweiligen Zielregionen benannt, heißen parasol-Ganglienzellen auch M-Zellen ("M" hier für magnozellulär) und midget-Ganglienzellen auch P-Zellen ("P" hier für parvozellulär); gelegentlich werden die bistratified-Ganglienzellen als K-Zellen ("K" für koniozellulär) bezeichnet.
Daneben finden sich im Säugetierauge auch 1–3 % an retinalen Ganglienzellen, die nicht zur visuellen Bildverarbeitung beitragen, sondern dem non-visuellen System okulärer Photosensitivität zugeordnet werden. Diese Neuronen enthalten ein Photopigment, Melanopsin, das sie selbst lichtempfindlich macht. Licht überführen sie in ein länger anhaltendes und depolarisiertes Rezeptorpotential, im Unterschied zu den ciliären Photorezeptoren mit Kontakt zum retinalen Pigmentepithel (RPE).
- (intrinsisch) photosensitive retinale Ganglienzellen (ipRGCs) – Hier werden bisher fünf Haupttypen (M1, M2, M3, M4, M5) mit Subtypen unterschieden. Doch sind deren Abgrenzungen noch unscharf und insbesondere ihre jeweilige physiologische Aufgabe im Einzelnen noch unklar. Sie erhalten aus der Retina Signale, vornehmlich von ON-Bipolaren Zellen, und geben auch Signale an Neuronen der Retina weiter, insbesondere Amakrinzellen. Ihre wesentliche Funktion wird jedoch getragen von ihren mit den Sehnerven verlaufenden Neuriten zu
- Kernen des Nucleus suprachiasmaticus im Hypothalamus (Licht als Zeitgeber circadianer Rhythmen),
- Kernen der Area pretectalis im Epithalamus (afferenter Reflexbogenanteil der Pupillenlichtreaktion),
- Kernen der oberen Hügel im Mittelhirn (Afferenzen für reflektorische Augenbewegungen).
Anmerkungen und Einzelnachweise
- Die Bezeichnung „Ganglienzelle“ geht zurück auf die anatomische Unterscheidung des Nervensystems in einen zentralen Anteil und den peripheren, bei dem dann Anhäufungen von Nervenzellkörpern als Ganglien bezeichnet werden. Derart wurden auch die Zellkörper von Neuronen der Netzhaut in der Inneren Körnerschicht (Stratum nucleare internum) als „Ganglion retinae“ und die in der Ganglienzellschicht (Stratum ganglionare) als „Ganglion nervi optici“ zusammengefasst. Nur letztere, die Sehnervenganglienzellen, werden üblicherweise „retinale Ganglienzellen“ (RGC) genannt.
- Eine knappe schematische Zusammenfassung nach Dacey et al.(Washington, 2003) wird wiedergegeben im Neuronbank Wiki unter Retinal Ganglion Cell, Types of RGCs (Memento des Originals vom 25. Juli 2009 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis..
- Eric R. Kandel, James H. Schwarz, Thomas M. Jessell (Hrsg.): Principles of neural science. 4. edition, international edition. McGraw-Hill, New York NY u. a. 2000, ISBN 0-8385-7701-6.
- Barry B. Lee, Paul R. Martin, Ulrike Grünert: Retinal connectivity and primate vision In: Progress in Retinal and Eye Research. Bd. 29, Nr. 6, November 2010, ISSN 1350-9462, S. 622–639, doi:10.1016/j.preteyeres.2010.08.004
- Gerald H. Jacobs, Jeremy Nathans: Der merkwürdige Farbensinn der Primaten. In: Spektrum der Wissenschaft. 5/2010, S. 44–51.